Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 9(6); 2018 > Article
-
Original Article
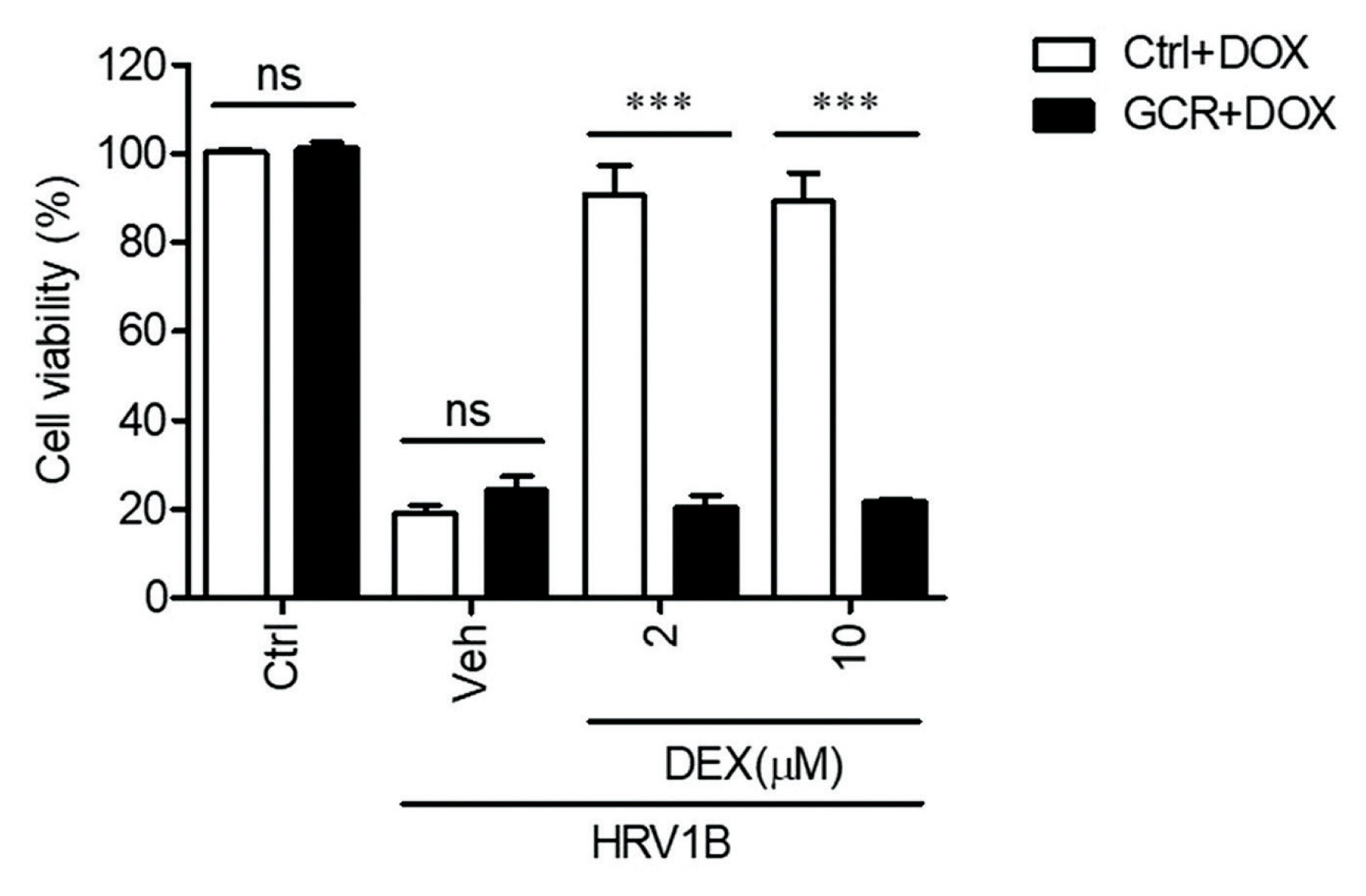
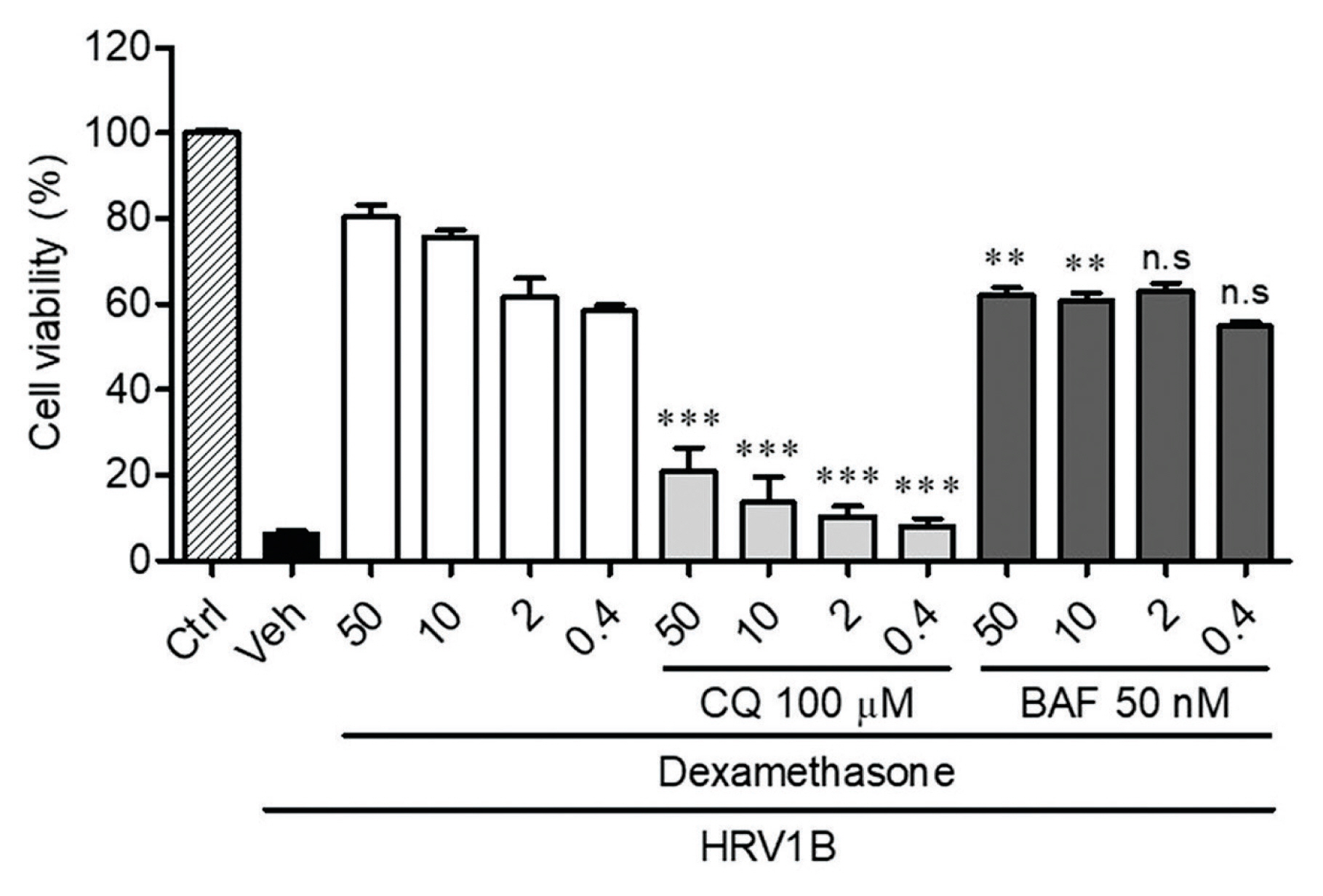
Anti-Human Rhinovirus 1B Activity of Dexamethasone viaGCR-Dependent Autophagy Activation - Jae-Sug Leea, Seong-Ryeol Kimb, Jae-Hyoung Songb, Yong-Pyo Leec, Hyun-Jeong Kob
-
Osong Public Health and Research Perspectives 2018;9(6):334-339.
DOI: https://doi.org/10.24171/j.phrp.2018.9.6.07
Published online: December 31, 2018
aDepartment of Beauty Science, Kwangju Women’s University, Gwangju, Korea
bLaboratory of Microbiology and Immunology, College of Pharmacy, Kangwon National University, Chuncheon, Korea
cDivision of Viral Diseases, Center for Laboratory Control and Infectious Diseases, Korea Centers for Disease Control and Prevention, Cheongju, Korea
- *Corresponding author: Hyun-Jeong Ko, Laboratory of Microbiology and Immunology, College of Pharmacy, Kangwon National University, Chuncheon, Korea, E-mail: hjko@kangwon.ac.kr
• Received: November 14, 2018 • Revised: November 21, 2018 • Accepted: November 21, 2018
Copyright ©2018, Korea Centers for Disease Control and Prevention
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
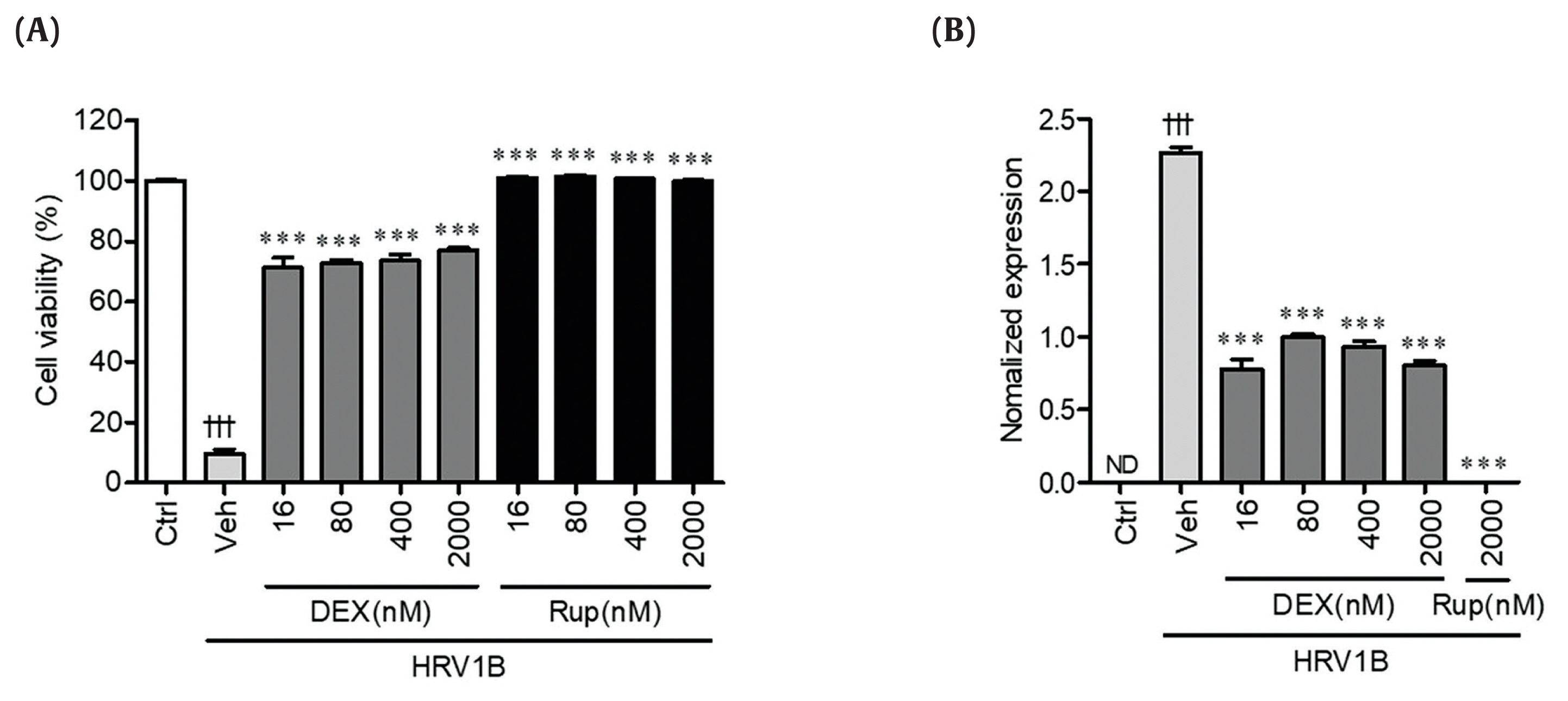
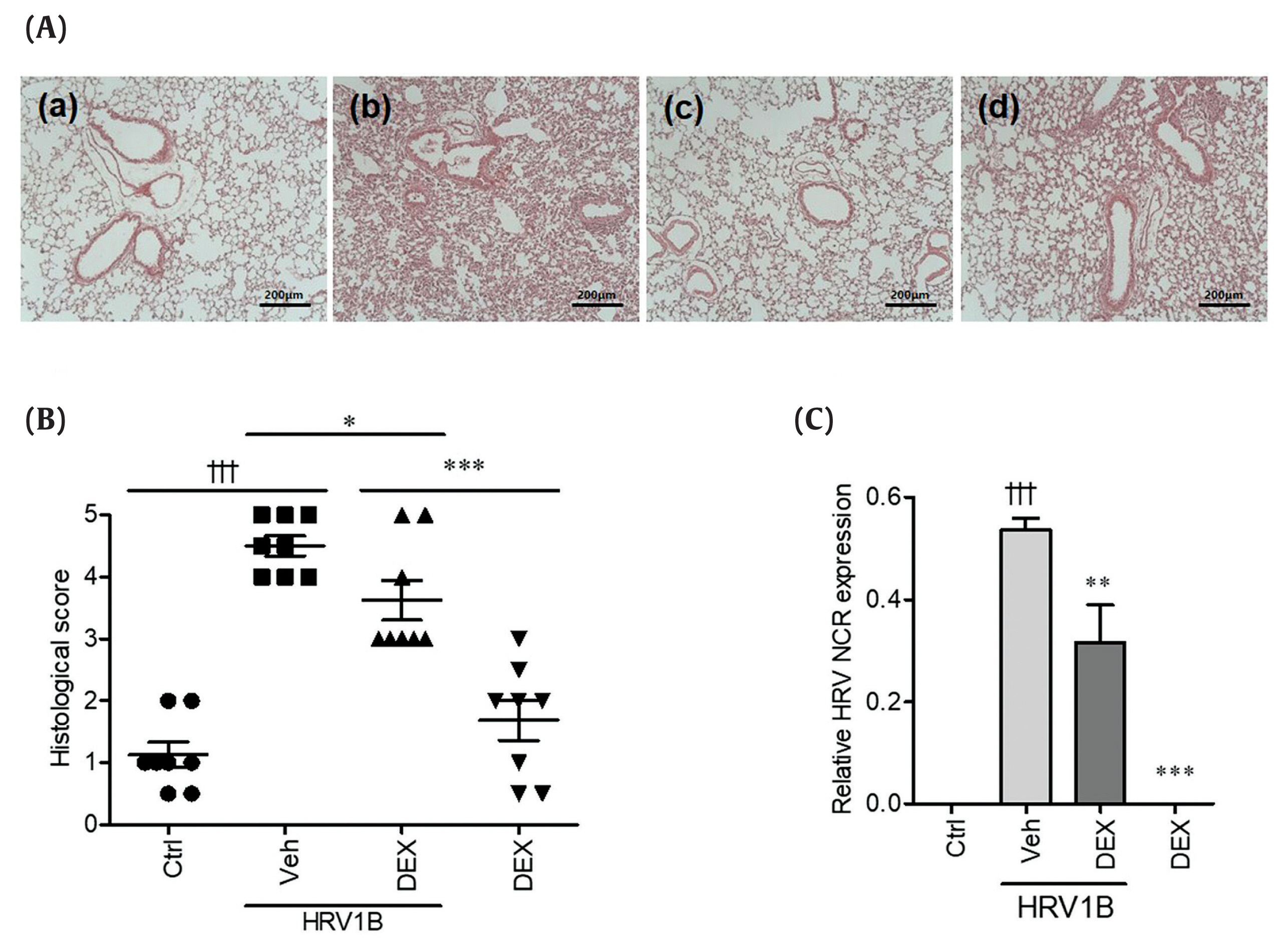


Figure & Data
References
Citations
Citations to this article as recorded by 
- Intracellular translocation of HMGB1 is important for Zika virus replication in Huh7 cells
Kim-Ling Chin, Nurhafiza Zainal, Sing-Sin Sam, Pouya Hassandarvish, Rafidah Lani, Sazaly AbuBakar
Scientific Reports.2022;[Epub] CrossRef - Pharmacological Modulators of Autophagy as a Potential Strategy for the Treatment of COVID-19
Gustavo José da Silva Pereira, Anderson Henrique França Figueredo Leão, Adolfo Garcia Erustes, Ingrid Beatriz de Melo Morais, Talita Aparecida de Moraes Vrechi, Lucas dos Santos Zamarioli, Cássia Arruda Souza Pereira, Laís de Oliveira Marchioro, Letícia P
International Journal of Molecular Sciences.2021; 22(8): 4067. CrossRef - MSTN Attenuates Cardiac Hypertrophy through Inhibition of Excessive Cardiac Autophagy by Blocking AMPK /mTOR and miR-128/PPARγ/NF-κB
Hanping Qi, Jing Ren, Lina Ba, Chao Song, Qianhui Zhang, Yonggang Cao, Pilong Shi, Bowen Fu, Yongsheng Liu, Hongli Sun
Molecular Therapy - Nucleic Acids.2020; 19: 507. CrossRef - Autophagy Augmentation to Alleviate Immune Response Dysfunction, and Resolve Respiratory and COVID-19 Exacerbations
Garrett Pehote, Neeraj Vij
Cells.2020; 9(9): 1952. CrossRef
 PubReader
PubReader Cite
Cite