
Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 8(5); 2017 > Article
-
Original Article
Antiviral Activity of Itraconazole against Echovirus 30 InfectionIn Vitro - Jae-Sug Leea, Hwa-Jung Choia, Jae-Hyoung Songb, Hyun-Jeong Kob, Kyungah Yoonc, Jeong-Min Seongd
-
Osong Public Health and Research Perspectives 2017;8(5):318-324.
DOI: https://doi.org/10.24171/j.phrp.2017.8.5.05
Published online: October 31, 2017
aDepartment of Beauty Science, Kwangju Women’s University, Gwangju, Korea
bLaboratory of Microbiology and Immunology, College of Pharmacy, Kangwon National University, Chuncheon, Korea
cDepartment of Clinical Pathology, Daejeon Health Institute of Technology, Daejeon, Korea
dDepartment of Dental Hygiene, College of Health Science, Kangwon National University, Samcheok, Korea
- Corresponding author: Kyungah Yoon, E-mail: kayun@hit.ac.kr. Jeong-Min Seong, E-mail: jmseong@kangwon.ac.kr
- Corresponding author: Kyungah Yoon, E-mail: kayun@hit.ac.kr. Jeong-Min Seong, E-mail: jmseong@kangwon.ac.kr
- *Jae-Sug Lee and Hwa-Jung Choi contributed equally to this work.
• Received: August 28, 2017 • Revised: September 18, 2017 • Accepted: September 20, 2017
Copyright ©2017, Korea Centers for Disease Control and Prevention
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
Abstract
-
Objectives
- Echovirus 30 is a major cause of meningitis in children and adults. The aim of this study was to investigate whether the antifungal drug itraconazole could exhibit antiviral activity against echovirus 30.
-
Methods
- The cytopathic effect and viral RNA levels were assessed in RD cells as indicators of viral replication. The effects of itraconazole were compared to those of two known antiviral drugs, rupintrivir and pleconaril. The time course and time-of-addition assays were used to approximate the time at which itraconazole exerts its activity in the viral cycle.
-
Results
- Itraconazole and rupintrivir demonstrated the greatest potency against echovirus 30, demonstrating concentration-dependent activity, whereas pleconaril showed no antiviral activity. Itraconazole did not directly inactivate echovirus 30 particles or impede viral uptake into RD cells, but did affect the initial stages of echovirus 30 infection through interference with viral replication.
-
Conclusion
- Itraconazole can be considered a lead candidate for the development of antiviral drugs against echovirus 30 that may be used during the early stages of echovirus 30 replication.
- Viruses are the leading cause of meningitis worldwide, with an estimated incidence of 12 to 19 cases per 100,000 people per year [1]. Among the causative viral agents of meningitis, non-polio enteroviruses, especially echovirus 30, are predominant [2]. Echovirus 30 belongs to the species Enterovirus B of the genus Enterovirus of the Picornaviridae family, and has high genetic variability [3]. The genetic diversity of echovirus 30 has been studied and the virus variants were phylogenetically divided into several genogroups [4]. Epidemics of echovirus 30 infection have been noted in long-term surveillance reports from the USA, and recent outbreaks have also occurred in South America, Central America, Asia, and Europe [5].
- Itraconazole is a triazole antifungal drug that inhibits the enzyme lanosterol 14-α-demethylase, which is important in ergosterol synthesis in fungi [6]. It possesses greater activity than fluconazole, is less expensive than newer triazoles, and unlike its predecessor, ketoconazole, exerts minimal effects on mammalian enzymes [7]. Consequently, itraconazole is frequently used by veterinarians for the treatment of fungal infections [8]. Itraconazole was previously reported to have an effect on human immunodeficiency virus (HIV)-associated talaromycosis [9] and showed therapeutic and prophylactic activity against human rhinovirus infection in a murine model [10]. However, the antiviral activity of itraconazole against echovirus 30 has not yet been reported.
- In this study, we have investigated whether itraconazole exerts antiviral activity against echovirus 30 in vitro. Furthermore, we assessed the action of itraconazole on echovirus 30 replication in more detail.
INTRODUCTION
- 1. Virus, cells, and reagents
- Echovirus 30 was obtained from ATCC (American Type Culture Collection, Manassas, VA, USA) and propagated through the infection of RD cells, a human rhabdomyosarcoma cell line. RD cells were maintained in minimal essential medium (MEM) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic-antimycotic solution. MEM, FBS, trypsin-ethylenediaminetetraacetic acid, and antibiotic-antimycotic solution were purchased from Gibco BRL (Invitrogen Life Technologies, Karlsruhe, Germany). The antiviral activity of itraconazole against echovirus 30 was identified by in vitro screening of the Screen-WellTM FDA Approved Drug Library V2 version 1.0 (BML-2843-0100; Enzo Life Sciences Inc., Farmingdale, NY, USA). Rupintrivir and pleconaril were purchased from Sigma-Aldrich (St. Louis, MO, USA).
- 2. Antiviral and cytotoxicity assays
- The antiviral activity was evaluated by the sulforhodamine (SRB) method based on the cytopathic effect (CPE) induced by viral infection, as previously reported [11]. One day before infection, 2 × 104 RD cells/well were seeded into a 96-well culture plate (Falcon; BD Biosciences, San Jose, CA, USA) and incubated at 37°C in an atmosphere of 5% CO2. The next day, the medium was changed to MEM supplemented with 1% FBS, and the diluted virus suspension at a 50% cell culture infective dose (CCID50), and the indicated concentrations of the compounds were added. The culture plate was incubated at 37°C in 5% CO2 for 2 days until the appropriate CPE was achieved. The cells were fixed with ice-cold 70% acetone for 30 minutes and stained with 0.4% (w/v) SRB (Sigma-Aldrich) in 1% acetic acid. The bound SRB was then solubilized with 10 mM Tris base and the absorbance was read at 562 nm by using a SpectraMax® i3 microplate reader (Molecular Devices, Palo Alto, CA, USA), with a reference absorbance at 650 nm. The results of the antiviral activity were presented as a percentage of the control.
- To evaluate the cytotoxicity, RD cells (2 × 104 cells/well) were seeded into 96-well culture plates. The following day, the medium was removed and the 96-well plates were replaced with media containing serially diluted compounds. After incubation for 2 days, cytotoxicity was evaluated by using the SRB assay. The culture medium was removed and washed with PBS, followed by the antiviral activity assay described above. The results were calculated as a percentage of the controls.
- To test the effect of itraconazole on the infectivity of echovirus 30 particles, echovirus 30 was preincubated with itraconazole, rupintrivir, or pleconaril for 1 hour at 4°C, and RD cells were infected with pretreated or untreated echovirus 30 for 1 hour at 37°C. The unbound virus was removed and the cells were washed twice with PBS and incubated in medium supplemented with or without itraconazole, rupintrivir, or pleconaril at 37°C. The antiviral activity was determined by the SRB assay after 2 days [12].
- 3. Time course experiment
- RD cells were infected with echovirus 30 at the CCID50, treated with itraconazole, rupintrivir, or pleconaril, and harvested at the indicated time points (1, 2, 4, 6, 8, 10, and 12 hours post-infection). The total RNA was isolated and the level of echovirus 30 RNA was analyzed by quantitative reverse transcription polymerase chain reaction (RT-qPCR).
- 4. Time-of-addition (TOA) assay
- The TOA assay was designed to determine the mechanism of action of itraconazole [13,14]. Itraconazole, rupintrivir, or pleconaril were added to the cells at 10 μg/mL either 1 hour before (−1 hour), during (0 hour), or after (1, 2, 4, and 8 hours) echovirus 30 infection. After 12-hour incubation, real-time PCR analysis was performed by using the Thunderbird® SYBR® qPCR Mix (Toyobo, Osaka, Japan).
- 5. Quantitative reverse transcription polymerase chain reaction (RT-qPCR)
- Total RNA was extracted by using the QIAamp® viral RNA Mini kit (Qiagen, Valencia, CA, USA) in accordance with the manufacturer’s instructions. For a 20-μL reverse transcription reaction, RNA was incubated for 60 minutes at 42°C with RNase inhibitor, murine Moloney leukemia virus reverse transcriptase, oligo(dT)15 primer, and dNTPs in 1× buffer (all from Promega, Madison, WI, USA) in accordance with the manufacturer’s instructions. For the qPCR analysis, the cDNA was amplified by using the CFX96TM Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) with Thunderbird® SYBR® qPCR Mix and the following primers: echovirus 30 5′-noncoding region (NCR)-sense, 5′-TCC TCC GGC CCC TGA ATG-3′ and 5′-NCR-antisense, 5′-GAA ACA CGG ACA CCC AAA G-3′ [15].
- 6. Statistical analyses
- To compare multiple groups, one-way analysis of variance (ANOVA) followed by Bonferroni’s multiple comparison test was computed by GraphPad Prism version 5 (GraphPad Software, La Jolla, CA, USA). Values of p < 0.05 were considered to be statistically significant at a 95% confidence interval.
MATERIALS AND METHODS
- 1. Itraconazole has antiviral activity against echovirus 30 in vitro
- It was recently reported that itraconazole exerted antiviral activity against enteroviruses, including coxsackievirus B3 and human rhinoviruses [10]. Here, we confirmed the antiviral activity of itraconazole against echovirus 30 through the assessment of its virus-induced CPE in RD cells by using the SRB assay. Echovirus 30 infection induced cell death in approximately 95% of RD cells; the treatment of the cells with 2 μM itraconazole significantly increased the cell viability to 90%. Furthermore, 10 μM and 50 μM itraconazole completely protected RD cells from echovirus 30-induced cytotoxicity. Rupintrivir, used as a positive control, showed strong antiviral activity against echovirus 30 at 0.08 μM, 0.4 μM, 2 μM, and 10 μM. However, pleconaril did not exhibit antiviral activity at the test concentrations (Figure 1A).
- We also assessed the cytotoxicity of itraconazole, rupintrivir, and pleconaril in RD cells. Itraconazole and rupintrivir did not induce cytotoxicity at 10 μM and 50 μM, but pleconaril produced substantial cytotoxicity at 50 μM concentrations (Figure 1B). Collectively, these results indicated that itraconazole exerted antiviral activity against echovirus 30 in vitro.
- Based on the results of the echovirus 30 cytotoxicity assays, we performed time-course experiments to analyze the mechanism of activity of itraconazole. Echovirus 30 viral RNA was analyzed by using RT-qPCR at various time points (i.e., 1, 2, 4, 6, 8, 10, and 12 hours) after 10-μM itraconazole treatment of echovirus 30-infected cells. We detected the viral RNA of echovirus 30 from 8 hours post-infection. Interestingly, itraconazole effectively inhibited the proliferation of echovirus 30, similar to the well-known effect of rupintrivir, which is an inhibitor of picornavirus 3C protease (Figure 2). Based on the results of these time course experiments, we hypothesized that itraconazole impacts upon a relatively early stage of the viral cycle. Therefore, it could target the entry, uncoating, translation, protease polyprotein processing, or replication. To investigate which step was affected by itraconazole, we adopted a TOA experiment, in which 10 μM itraconazole was applied to the culture medium at −1, 0, 1, 2, 4, 6, 8, or 10 hours post-infection, and echovirus 30 RNA replication was analyzed at 12 hours post-infection.
- Itraconazole was added at different periods (before, during, and after) of echovirus 30 infection. The treatment of RD cells with itraconazole 1 hour before echovirus 30 infection did not inhibit the virus. However, the treatment with itraconazole during the 6-hour period after viral infection effectively suppressed viral RNA production. In contrast, the treatment with itraconazole at 8 hours after infection did not inhibit the virus effectively. The inhibitory effect of pleconaril on viral RNA production was not observed when added prior (−1 hour) or during (0 hour) or post (1, 2, 4, 6, 8, and 10 hours) infection. Rupintrivir effectively suppressed viral RNA production when added prior (−1 hour) or during (0 hour) or post (1, 2, 4, and 6 hours) infection and showed an excellent inhibitory effect when added at 8 and 10 hours post-infection (Figure 3). The effect of itraconazole and rupintrivir on the suppression of viral RNA production showed similar patterns (Figure 3). These results suggested that itraconazole did not suppress the entry of the virus into the cell, but could inhibit a critical step that occurs approximately 6 hours after infection (Figure 3).
- 2. Effects of itraconazole on the infectivity of echovirus 30 particles
- Next, we assessed the effects of itraconazole directly on the infectivity of the echovirus 30 particles. The preincubation of viral particles (−1 hour) with itraconazole, rupintrivir, or pleconaril did not significantly alter infectivity. In contrast, the continuous presence (from 0 hour) of itraconazole or rupintrivir during infection resulted in a marked increase in antiviral activity. However, pleconaril did not exhibit antiviral activity during infection (Figure 4). This result suggested that itraconazole does not bind and neutralize echovirus 30 particles because pre-exposure of the virus to itraconazole did not alter viral infectivity.
RESULTS
- The Enterovirus genus, Enterovirus B species, and Picornaviridae family include echovirus type 30, which is a single-stranded RNA virus. From asymptomatic infection, nonspecific febrile illness, nonspecific rash, and upper respiratory infections to sepsis, aseptic meningitis, encephalitis, and myositis, especially in neonates and children, echovirus is associated with a wide range of diseases. Echovirus replicates in the gastrointestinal tract. In addition, it is transmitted between individuals usually by the fecal-oral route, although transmission may occur through respiration [16–19].
- Echovirus 30 is one of the most frequently isolated echovirus serotypes that causes aseptic meningitis [20]. Numerous aseptic meningitis outbreaks that are caused by different strains of echovirus 30 have been reported in the last twenty years in various countries [21–23]. In the USA, the primary cause of meningitis outbreaks between 2003 and 2004 was echovirus 30 [4,24]. Furthermore, other outbreaks of echovirus 30-associated meningitis occurred in Asia in Taiwan (2001), China (2003 and 2004), and Japan (2004 and 2006) [25–28]. In Korea, human enterovirus (HEV)-associated meningitis outbreaks have been reported since 1993; echovirus 30 was responsible for an aseptic meningitis outbreak in 1997 [29]. Therefore, there is a need to identify inhibitors of echovirus 30 infections that may be developed for treatment.
- The triazole group of antifungals, of which itraconazole is a member, has an extensive spectrum of antifungal activity against various pathogenic fungi and yeasts, with low toxicity [30,31]. Thus, infection with Cryptococcus neoformans, Histoplasma capsulatum, and several other opportunistic fungal pathogens are treated with itraconazole in the form of an orally administered triazole antifungal drug. Itraconazole has also been used as a nasal spray for the cure of fungal infections. Furthermore, the antiviral effect of itraconazole has been demonstrated against HIV [32–34] and hepatitis [35–37]. However, to the best of our knowledge, there have been no previous definitive studies on the antiviral activity of itraconazole against echovirus 30. In the current study, we clarified the antiviral activity of itraconazole against echovirus 30 in RD cells. Treatment with 2 μM itraconazole significantly reduced echovirus 30 RNA levels by 90% and exhibited complete antiviral activity against echovirus 30 at concentrations over 10 μM, but no cytotoxicity was observed at all tested concentrations. Therefore, itraconazole, which exhibited potent inhibitory effects on viral production, represents a novel candidate for echovirus 30 therapy.
- Pleconaril is a picornavirus capsid binding inhibitor that prevents attachment and/or virus uncoating [38]. Rupintrivir is a rhinovirus 3C protease inhibitor, which reached phase II clinical trials in 1999 [39]. The two compounds were evaluated against echovirus 30 in CPE assays to determine viral inhibition. Pleconaril did not show antiviral activity, but 0.08 μM rupintrivir, which was used as the positive control, showed strong antiviral activity against echovirus 30. Rupintrivir (10 μM) was not cytotoxic in RD cells, but addition of 50 μM pleconaril resulted in high cytotoxicity. Therefore, our results indicated that rupintrivir is active against echovirus 30 in RD cells.
- To investigate the action of itraconazole on echovirus 30 in more detail, we investigated the effect of the drug in multiple steps of the viral infection cycle. Up to 6 hours post-infection, itraconazole effectively inhibited the expression of echovirus 30 RNA in a similar manner to the positive control, rupintrivir. In a TOA experiment, the treatment with itraconazole 1 hour prior- and 8 hours post-infection did not inhibit the expression of echovirus 30 RNA, but treatment with itraconazole between 0 and 4 hours post-infection effectively suppressed expression of echovirus 30 RNA. The pre-exposure of the virus to itraconazole, prior to RD cell infection, did not alter the infectivity of the echovirus 30 particles.
- Collectively, itraconazole did not suppress the entry of the virus into the cell, but was able to inhibit the early stages of the viral cycle that occur after viral entry. Our results provided a new direction for the development of potential antiviral agents against echovirus 30 infection.
DISCUSSION
-
Acknowledgements
- This study was supported by a 2015 Research Grant from Kangwon National University. This research was also supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT, and Future Planning (NRF-2016R1A6A3A11932323). In addition, this study was supported (in part) by Research Funds of Kwangju Women’s University (Gwangju, Korea) in 2017 (KWUI17-032).
ACKNOWLEDGMENTS
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
Article information
- 1. Rudolph H, Prieto Dernbach R, Walka M, et al. Comparison of clinical and laboratory characteristics during two major paediatric meningitis outbreaks of echovirus 30 and other non-polio enteroviruses in Germany in 2008 and 2013. Eur J Clin Microbiol Infect Dis 2017;In press. https://doi.org/10.1007/s10096-017-2979-7. PMID: 10.1007/s10096-017-2979-7. PMID: 28409290.Article
- 2. Rudolph H, Schroten H, Tenenbaum T. Enterovirus infections of the central nervous system in children: an update. Pediatr Infect Dis J 2016;35:567−9. https://doi.org/10.1097/INF.0000000000001090. PMID: 10.1097/INF.0000000000001090. PMID: 26862675.ArticlePubMed
- 3. Wieczorek M, Krzysztoszek A, Figas A. Molecular characterization of echovirus 30 isolates from Poland, 1995–2015. Virus Genes 2016;52:400−4. https://doi.org/10.1007/s11262-016-1310-5. PMID: 10.1007/s11262-016-1310-5. PMID: 26957092.ArticlePubMed
- 4. Oberste MS, Maher K, Kennett ML, et al. Molecular epidemiology and genetic diversity of echovirus type 30 (E30): genotypes correlate with temporal dynamics of E30 isolation. J Clin Microbiol 1999;37:3928−33. PMID: 10565909.ArticlePubMedPMC
- 5. Österback R, Kalliokoski T, Lähdesmäki T, et al. Echovirus 30 meningitis epidemic followed by an outbreak-specific RT-qPCR. J Clin Virol 2015;69:7−11. https://doi.org/10.1016/j.jcv.2015.05.012. PMID: 10.1016/j.jcv.2015.05.012. PMID: 26209368.ArticlePubMed
- 6. Bae JH, Hwang AR, Kim CY, et al. Intravitreal itraconazole inhibits laser-induced choroidal neovascularization in rats. PLoS One 2017;12:e0180482https://doi.org/10.1371/journal.pone.0180482. PMID: 10.1371/journal.pone.0180482. PMID: 28666022.ArticlePubMedPMC
- 7. Lestner J, Hope WW. Itraconazole: an update on pharmacology and clinical use for treatment of invasive and allergic fungal infections. Expert Opin Drug Metab Toxicol 2013;9:911−26. https://doi.org/10.1517/17425255.2013.794785. PMID: 10.1517/17425255.2013.794785. PMID: 23641752.ArticlePubMed
- 8. Hasbach AE, Langlois DK, Rosser EJ Jr, et al. Pharmacokinetics and relative bioavailability of orally administered innovator-formulated itraconazole capsules and solution in healthy dogs. J Vet Intern Med 2017;31:1163−9. https://doi.org/10.1111/jvim.14779. PMID: 10.1111/jvim.14779. PMID: 28627123.ArticlePubMedPMC
- 9. Le T, Kinh NV, Cuc NTK, et al. A trial of itraconazole or amphotericin B for HIV-associated talaromycosis. N Engl J Med 2017;376:2329−40. https://doi.org/10.1056/NEJMoa1613306. PMID: 10.1056/NEJMoa1613306. PMID: 28614691.ArticlePubMed
- 10. Shim A, Song JH, Kwon BE, et al. Therapeutic and prophylactic activity of itraconazole against human rhinovirus infection in a murine model. Sci Rep 2016;6:23110https://doi.org/10.1038/srep23110. PMID: 10.1038/srep23110. PMID: 26976677.ArticlePubMedPMC
- 11. Choi HJ, Song HH, Lee JS, et al. Inhibitory effects of norwogonin, oroxylin A, and mosloflavone on enterovirus 71. Biomol Ther (Seoul) 2016;24:552−8. https://doi.org/10.4062/biomolther.2015.200. PMID: 10.4062/biomolther.2015.200. PMID: 27257010.ArticlePubMedPMC
- 12. Choi HJ, Kim JH, Lee CH, et al. Antiviral activity of quercetin 7-rhamnoside against porcine epidemic diarrhea virus. Antiviral Res 2009;81:77−81. https://doi.org/10.1016/j.antiviral.2008.10.002. PMID: 10.1016/j.antiviral.2008.10.002. PMID: 18992773.ArticlePubMed
- 13. Daelemans D, Pauwels R, De Clercq E, et al. A time-of-drug addition approach to target identification of antiviral compounds. Nat Protoc 2011;6:925−33. https://doi.org/10.1038/nprot.2011.330. PMID: 10.1038/nprot.2011.330. PMID: 21637207.ArticlePubMedPMC
- 14. Basu A, Li B, Mills DM, et al. Identification of a small-molecule entry inhibitor for filoviruses. J Virol 2011;85:3106−19. https://doi.org/10.1128/JVI.01456-10. PMID: 10.1128/JVI.01456-10. PMID: 21270170.ArticlePubMedPMC
- 15. Kwon BE, Song JH, Song HH, et al. Antiviral activity of oroxylin A against coxsackievirus B3 alleviates virus-induced acute pancreatic damage in mice. PLoS One 2016;11:e0155784https://doi.org/10.1371/journal.pone.0155784. PMID: 10.1371/journal.pone.0155784. PMID: 27195463.ArticlePubMedPMC
- 16. Harvala H, Simmonds P. Human parechoviruses: biology, epidemiology and clinical significance. J Clin Virol 2009;45:1−9. https://doi.org/10.1016/j.jcv.2009.03.009. PMID: 10.1016/j.jcv.2009.03.009. PMID: 19372062.ArticlePubMed
- 17. Esposito S, Rahamat-Langendoen J, Ascolese B, et al. Pediatric parechovirus infections. J Clin Virol 2014;60:84−9. https://doi.org/10.1016/j.jcv.2014.03.003. PMID: 10.1016/j.jcv.2014.03.003. PMID: 24690382.ArticlePubMed
- 18. Wolthers KC, Benschop KS, Schinkel J, et al. Human parechoviruses as an important viral cause of sepsislike illness and meningitis in young children. Clin Infect Dis 2008;47:358−63. https://doi.org/10.1086/589752. PMID: 10.1086/589752. PMID: 18558876.ArticlePubMed
- 19. Lee JY, Seo Y, Choi UY, et al. Seroepidemiology of echovirus 30 in Korean children. World J Pediatr 2017;In press. https://doi.org/10.1007/s12519-017-0058-x. PMID: 10.1007/s12519-017-0058-x. PMID: 28766163.Article
- 20. Lu J, Zheng H, Guo X, et al. Elucidation of echovirus 30’s origin and transmission during the 2012 aseptic meningitis outbreak in Guangdong, China, through continuing environmental surveillance. Appl Environ Microbiol 2015;81:2311−9. https://doi.org/10.1128/AEM.03200-14. PMID: 10.1128/AEM.03200-14. PMID: 25616804.ArticlePubMedPMC
- 21. dos Santos GP, da Costa EV, Tavares FN, et al. Genetic diversity of Echovirus 30 involved in aseptic meningitis cases in Brazil (1998–2008). J Med Virol 2011;83:2164−71. https://doi.org/10.1002/jmv.22235. PMID: 10.1002/jmv.22235. PMID: 22012725.ArticlePubMed
- 22. Xiao H, Guan D, Chen R, et al. Molecular characterization of echovirus 30-associated outbreak of aseptic meningitis in Guangdong in 2012. Virol J 2013;10:263https://doi.org/10.1186/1743-422X-10-263. PMID: 10.1186/1743-422X-10-263. PMID: 23968330.ArticlePubMedPMC
- 23. Milia MG, Cerutti F, Gregori G, et al. Recent outbreak of aseptic meningitis in Italy due to Echovirus 30 and phylogenetic relationship with other European circulating strains. J Clin Virol 2013;58:579−83. https://doi.org/10.1016/j.jcv.2013.08.023. PMID: 10.1016/j.jcv.2013.08.023. PMID: 24051044.ArticlePubMed
- 24. Mirand A, Henquell C, Archimbaud C, et al. Emergence of recent echovirus 30 lineages is marked by serial genetic recombination events. J Gen Virol 2007;88(Pt 1). 166−76. https://doi.org/10.1099/vir.0.82146-0. PMID: 10.1099/vir.0.82146-0. PMID: 17170449.ArticlePubMed
- 25. Akiyoshi K, Nakagawa N, Suga T. An outbreak of aseptic meningitis in a nursery school caused by echovirus type 30 in Kobe, Japan. Jpn J Infect Dis 2007;60:66−8. PMID: 17314434.ArticlePubMed
- 26. Wang JR, Tsai HP, Huang SW, et al. Laboratory diagnosis and genetic analysis of an echovirus 30-associated outbreak of aseptic meningitis in Taiwan in 2001. J Clin Microbiol 2002;40:4439−44. https://doi.org/10.1128/JCM.40.12.4439-4444.2002. PMID: 10.1128/JCM.40.12.4439-4444.2002. PMID: 12454133.ArticlePubMedPMC
- 27. Wang HY, Xu AQ, Zhu Z, et al. The genetic characterization and molecular evolution of echovirus 30 during outbreaks of aseptic meningitis. Zhonghua Liu Xing Bing Xue Za Zhi 2006;27:793−7. In Chinese. https://doi.org/10.3760/cma.j.issn.2095-1485.2006.09.001. PMID: 17299967.
- 28. Zhao YN, Jiang QW, Jiang RJ, et al. Echovirus 30, Jiangsu Province, China. Emerg Infect Dis 2005;11:562−7. https://doi.org/10.3201/eid1104.040995. PMID: 10.3201/eid1104.040995. PMID: 15829194.ArticlePubMedPMC
- 29. Choi YJ, Park KS, Baek KA, et al. Molecular characterization of echovirus 30-associated outbreak of aseptic meningitis in Korea in 2008. J Microbiol Biotechnol 2010;20:643−9. PMID: 20372040.PubMed
- 30. Sozen H, Celik OI, Cetin ES, et al. Evaluation of the protective effect of silibinin in rats with liver damage caused by itraconazole. Cell Biochem Biophys 2015;71:1215−23. https://doi.org/10.1007/s12013-014-0331-8. PMID: 10.1007/s12013-014-0331-8. PMID: 25395192.ArticlePubMed
- 31. Saag MS, Dismukes WE. Azole antifungal agents: emphasis on new triazoles. Antimicrob Agents Chemother 1988;32:1−8. https://doi.org/10.1128/AAC.32.1.1. PMID: 10.1128/AAC.32.1.1. PMID: 2831809.ArticlePubMedPMC
- 32. McKinsey DS, Wheat LJ, Cloud GA, et al. Itraconazole prophylaxis for fungal infections in patients with advanced human immunodeficiency virus infection: randomized, placebo-controlled, double-blind study. National Institute of Allergy and Infectious Diseases Mycoses Study Group. Clin Infect Dis 1999;28:1049−56. https://doi.org/10.1086/514744. PMID: 10.1086/514744. PMID: 10452633.ArticlePubMed
- 33. Graybill JR, Vazquez J, Darouiche RO, et al. Randomized trial of itraconazole oral solution for oropharyngeal candidiasis in HIV/AIDS patients. Am J Med 1998;104:33−9. https://doi.org/10.1016/S0002-9343(97)00307-0. PMID: 10.1016/S0002-9343(97)00307-0. PMID: 9528717.ArticlePubMed
- 34. Murray PA, Koletar SL, Mallegol I, et al. Itraconazole oral solution versus clotrimazole troches for the treatment of oropharyngeal candidiasis in immunocompromised patients. Clin Ther 1997;19:471−80. https://doi.org/10.1016/S0149-2918(97)80131-2. PMID: 10.1016/S0149-2918(97)80131-2. PMID: 9220211.ArticlePubMed
- 35. Tuccori M, Bresci F, Guidi B, et al. Fatal hepatitis after long-term pulse itraconazole treatment for onychomycosis. Ann Pharmacother 2008;42:1112−7. https://doi.org/10.1345/aph.1L051. PMID: 10.1345/aph.1L051. PMID: 18523232.ArticlePubMed
- 36. Wang HF, Su HB, Li HW, et al. Severe viral hepatitis complicated with pleural aspergillus infection treated with itraconazole and amphotericin B. Zhonghua Gan Zang Bing Za Zhi 2006;14:690−1. In Chinese. https://doi.org/10.3760/cma.j.issn.1004-6461.2006.09.019. PMID: 16995989.
- 37. Adriaenssens B, Roskams T, Steger P, et al. Hepatotoxicity related to itraconazole: report of three cases. Acta Clin Belg 2001;56:364−9. https://doi.org/10.1179/acb.2001.055. PMID: 10.1179/acb.2001.055. PMID: 11881322.ArticlePubMed
- 38. Smee DF, Evans WJ, Nicolaou KC, et al. Susceptibilities of enterovirus D68, enterovirus 71, and rhinovirus 87 strains to various antiviral compounds. Antiviral Res 2016;131:61−5. https://doi.org/10.1016/j.antiviral.2016.04.003. PMID: 10.1016/j.antiviral.2016.04.003. PMID: 27063860.ArticlePubMedPMC
- 39. Hayden FG, Turner RB, Gwaltney JM, et al. Phase II, randomized, double-blind, placebo-controlled studies of ruprintrivir nasal spray 2-percent suspension for prevention and treatment of experimentally induced rhinovirus colds in healthy volunteers. Antimicrob Agents Chemother 2003;47:3907−16. https://doi.org/10.1128/AAC.47.12.3907-3916.2003. PMID: 10.1128/AAC.47.12.3907-3916.2003. PMID: 14638501.ArticlePubMedPMC
REFERENCES
Figure 1
Itraconazole exerts antiviral activity against echovirus 30 in vitro. (A) The antiviral activity and (B) cytotoxicity of itraconazole were evaluated based on the cell viability of RD cells. The cell viability was evaluated by using the sulforhodamine assay and the results were determined based on the absorbance at 562 nm. The bar graphs show the mean ± standard deviation.
Ctrl, control; Veh, vehicle.

Figure 2The time course of echovirus 30 infection. RD cells infected with the 50% cell culture infective dose of echovirus 30 were harvested at the indicated time points after 2 μM itraconazole, 2 μM rupintrivir, or 10 μM pleconaril had been added (i.e., post-infection). The total RNA was isolated and echovirus 30 RNA was analyzed by RT-qPCR.

Figure 3Time-of-addition experiment testing the effect of itraconazole on the echovirus 30 viral cycle. Itraconazole (2 μM), rupintrivir (2 μM), or pleconaril (10 μM) were added prior to, at the time of, or after viral infection of RD cells, specifically at the indicated time points. The percentage of viable cells was analyzed 14 hours post-infection. RD cells that were treated with drugs prior to viral infection were washed before infection.
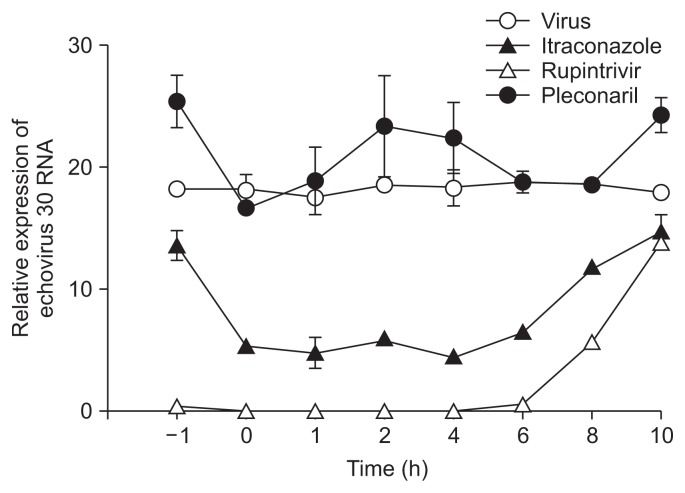
Figure 4Effects of itraconazole on infectivity of echovirus 30 particles. Echovirus 30 particles were incubated with of 2 μM itraconazole, 2 μM rupintrivir, or 10 μM pleconaril for 1 hour at 4°C. RD cells were then incubated in the presence or absence of virus for 1 hour at 37°C. The unbound virus was removed by extensive washing and the incubation was continued with or without 2 μM itraconazole, 2 μM rupintrivir, or 10 μM pleconaril at 37°C. The antiviral activity was determined by RT-qPCR analysis 2 days after infection. −1 h, pre-incubation of virus with the indicated drug without subsequent drug treatment of the infected cells; 0 h, incubation of cells with the indicated drug after viral infection; Ctrl, control; Veh, vehicle.
Figure & Data
References
Citations
Citations to this article as recorded by 
- Direct-Acting Antivirals and Host-Targeting Approaches against Enterovirus B Infections: Recent Advances
Chiara Tammaro, Michela Guida, Federico Appetecchia, Mariangela Biava, Sara Consalvi, Giovanna Poce
Pharmaceuticals.2023; 16(2): 203. CrossRef - Antiviral Activity of Approved Antibacterial, Antifungal, Antiprotozoal and Anthelmintic Drugs: Chances for Drug Repurposing for Antiviral Drug Discovery
Leena Abdulaziz, Esraa Elhadi, Ejlal A Abdallah, Fadlalbaseer A Alnoor, Bashir A Yousef
Journal of Experimental Pharmacology.2022; Volume 14: 97. CrossRef - Identification of novel Ebola virus inhibitors using biologically contained virus
Bert Vanmechelen, Joren Stroobants, Winston Chiu, Joost Schepers, Arnaud Marchand, Patrick Chaltin, Kurt Vermeire, Piet Maes
Antiviral Research.2022; 200: 105294. CrossRef - The Antifungal Itraconazole Is a Potent Inhibitor of Chikungunya Virus Replication
Lucca Policastro, Isabela Dolci, Andre Godoy, José Silva Júnior, Uriel Ruiz, Igor Santos, Ana Jardim, Kirandeep Samby, Jeremy Burrows, Timothy Wells, Laura Gil, Glaucius Oliva, Rafaela Fernandes
Viruses.2022; 14(7): 1351. CrossRef - Antifungal Triazole Posaconazole Targets an Early Stage of the Parechovirus A3 Life Cycle
Eric Rhoden, Terry Fei Fan Ng, Ray Campagnoli, W. Allan Nix, Jennifer Konopka-Anstadt, Rangaraj Selvarangan, Laurence Briesach, M. Steven Oberste, William C. Weldon
Antimicrobial Agents and Chemotherapy.2020;[Epub] CrossRef - Potential antiviral properties of antifungal drugs
FalahH.O Al-Khikani, HudaA.S Almosawey, YounusJ Abdullah, AtyafA Al-Asadi, RaghdahM Hameed, NoorF Hasan, MohanadK.M Al-Ibraheemi
Journal of the Egyptian Women's Dermatologic Socie.2020; 17(3): 185. CrossRef - Repurposing approach identifies new treatment options for invasive fungal disease
Isis Regina Grenier Capoci, Daniella Renata Faria, Karina Mayumi Sakita, Franciele Abigail Vilugron Rodrigues-Vendramini, Patricia de Souza Bonfim-Mendonça, Tania Cristina Alexandrino Becker, Érika Seki Kioshima, Terezinha Inez Estivalet Svidzinski, Berna
Bioorganic Chemistry.2019; 84: 87. CrossRef
 PubReader
PubReader Cite
Cite



