Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 11(4); 2020 > Article
-
Original Article
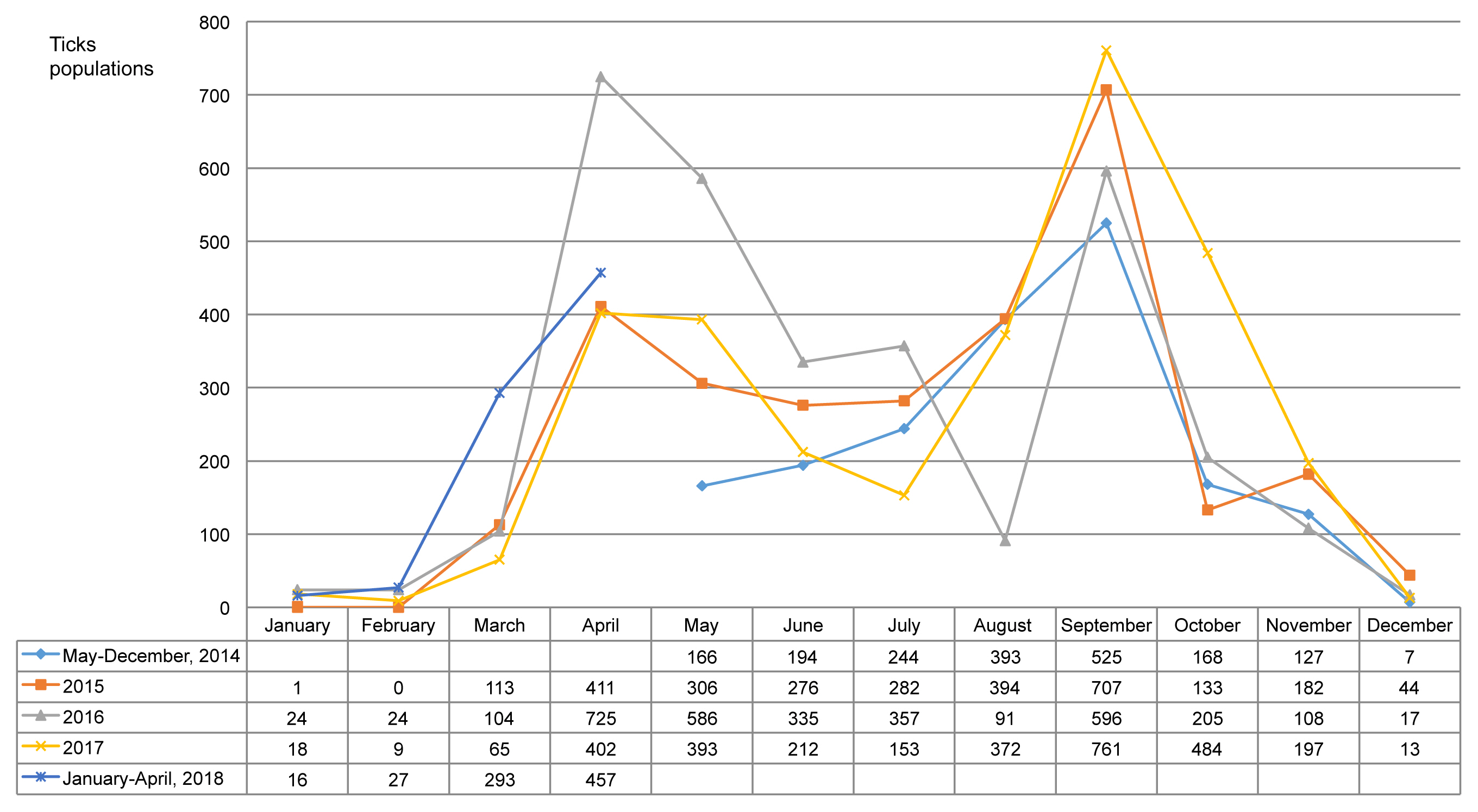
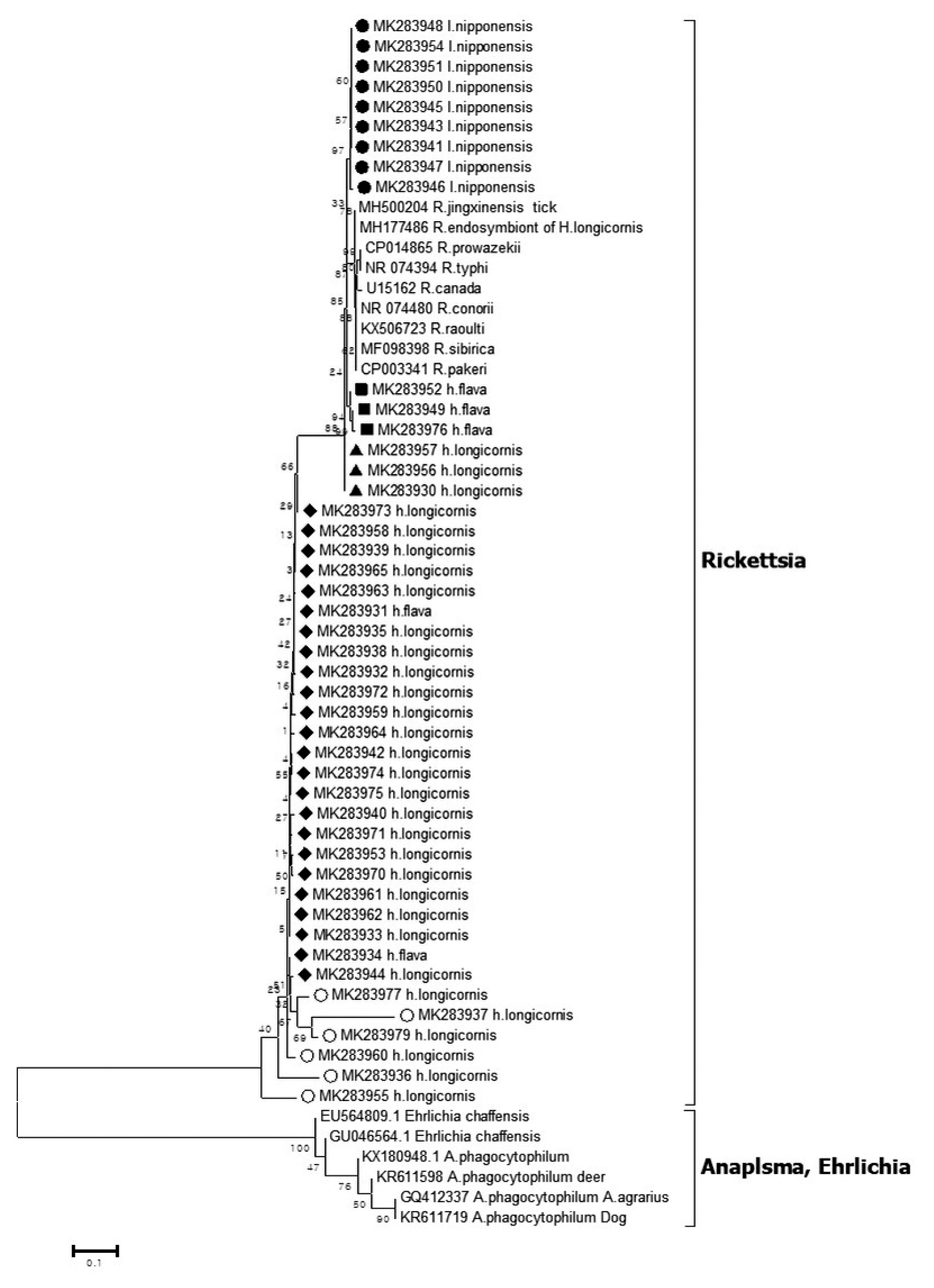
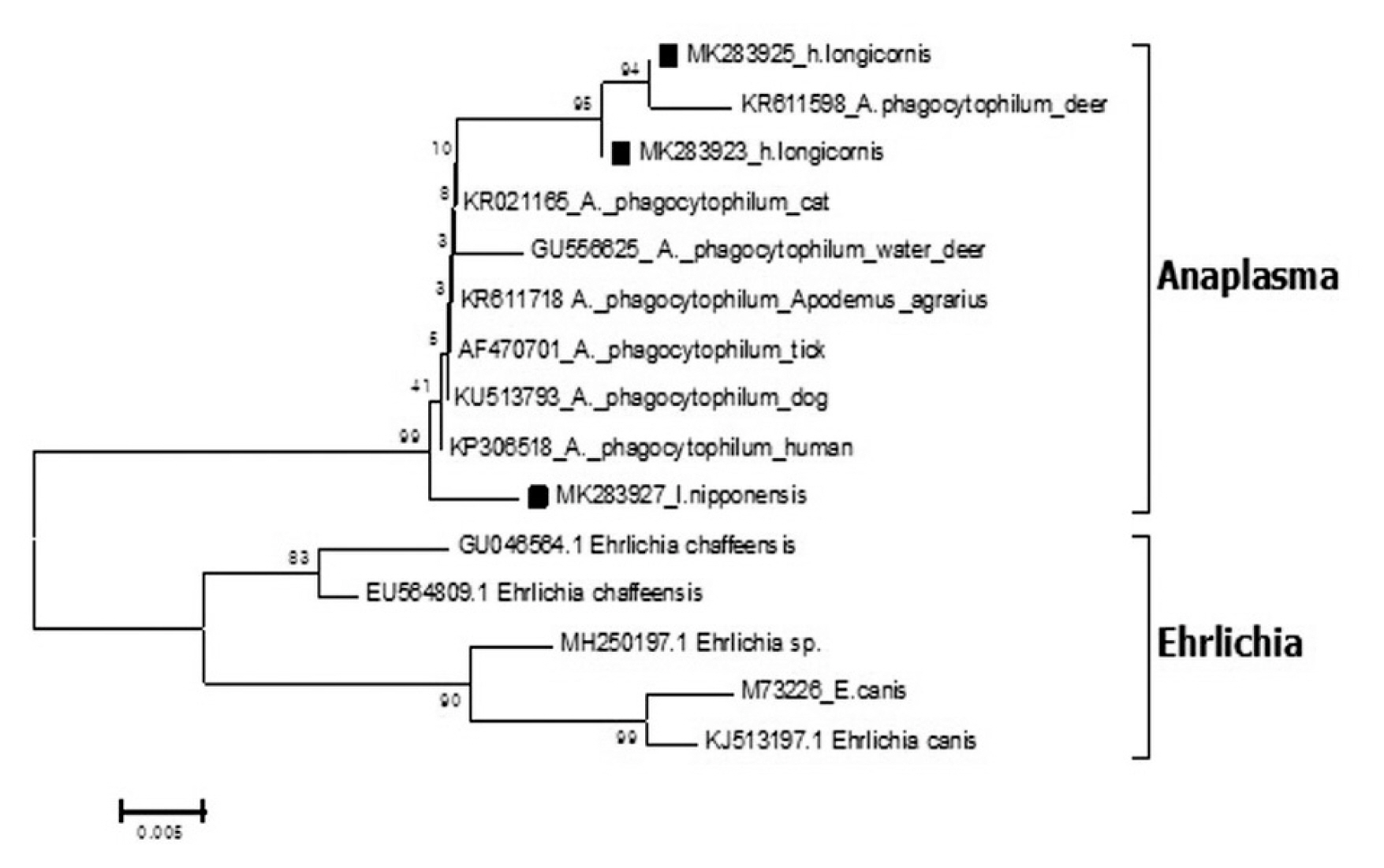
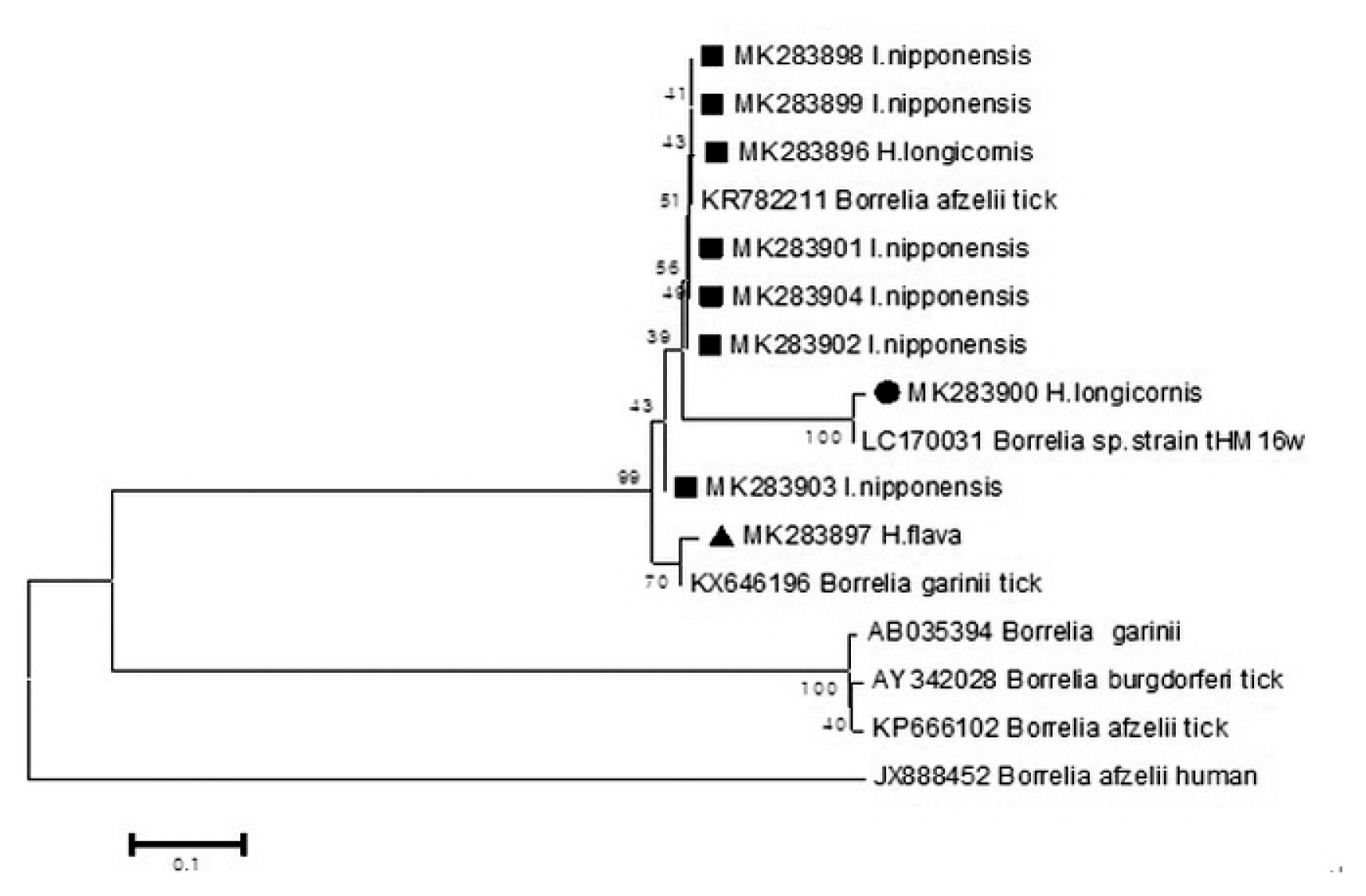
Epidemiological Characteristics of Field Tick-Borne Pathogens in Gwang-ju Metropolitan Area, South Korea, from 2014 to 2018 - Jung Wook Parka, Seung Hun Leeb,c, Gi Seong Leea, Jin Jong Seoa, Jae Keun Chunga
-
Osong Public Health and Research Perspectives 2020;11(4):177-184.
DOI: https://doi.org/10.24171/j.phrp.2020.11.4.06
Published online: August 31, 2020
aDivision of Infectious Disease Investigation, Health and Environment Research Institute of Gwangju, Gwangju, Korea
bDivision of Bacterial Disease Research, Center for Infectious Disease Research, Korea National Institute of Health, Cheongju, Korea
cYeosu National Quarantine Station, Korea Centers for Disease Control & Prevention, Yeosu, Korea
- *Corresponding author: Jae Keun Chung, Division of Infectious Disease Investigation, Health and Environment Research Institute of Gwangju, Gwangju 61986, Korea, E-mail: jkchung@korea.kr
• Received: December 26, 2019 • Revised: April 6, 2020 • Accepted: May 15, 2020
Copyright ©2020, Korea Centers for Disease Control and Prevention
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).

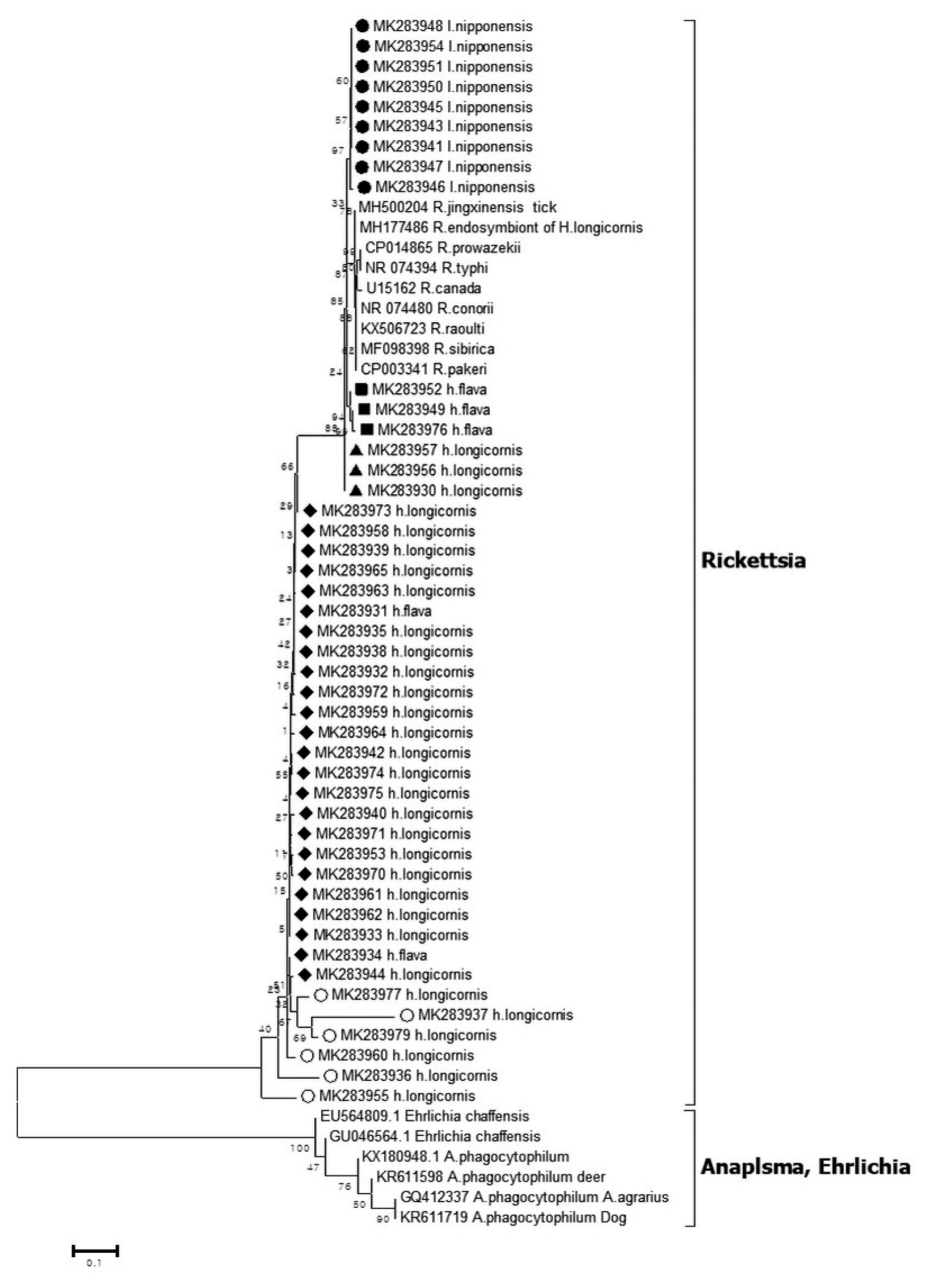
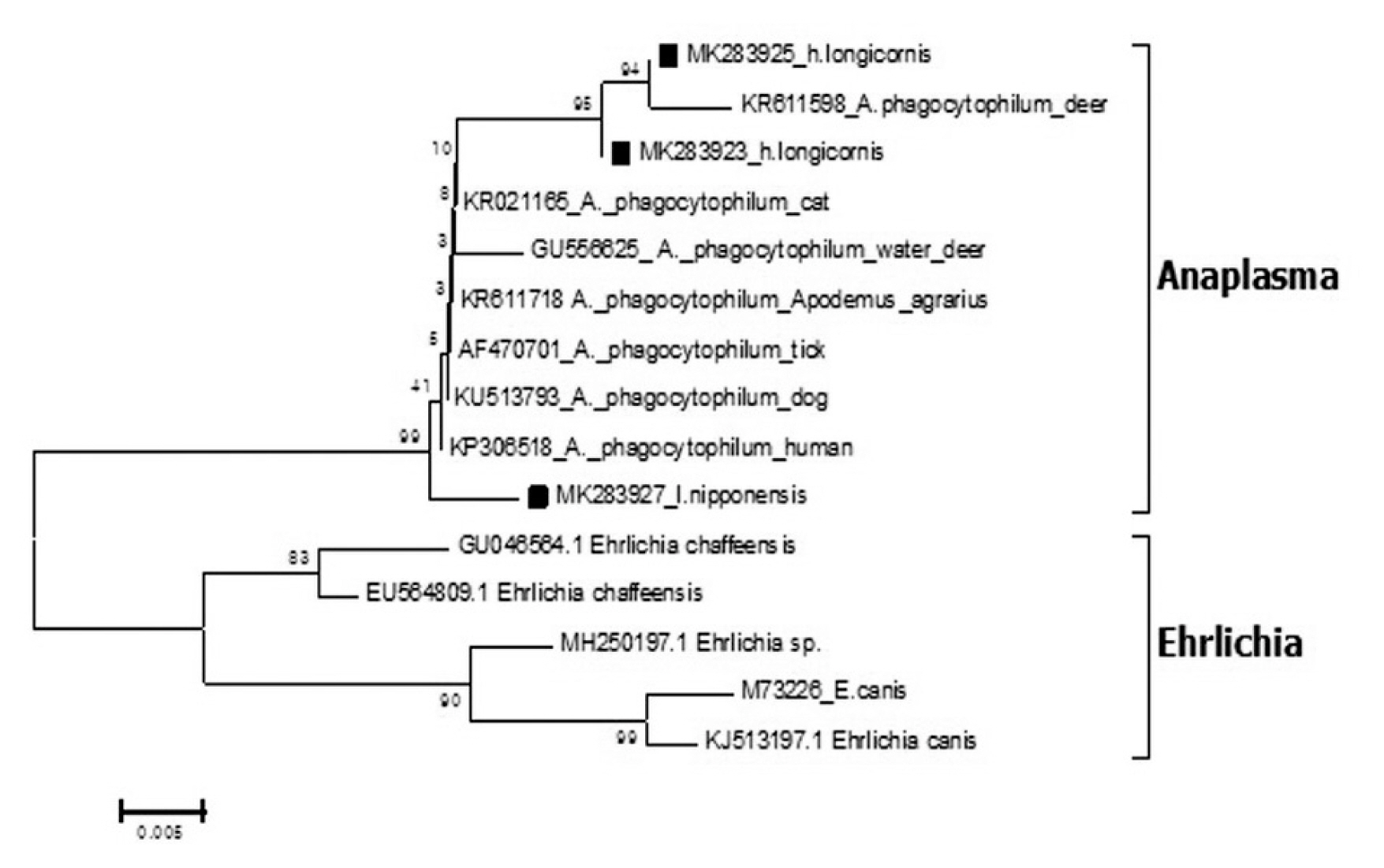
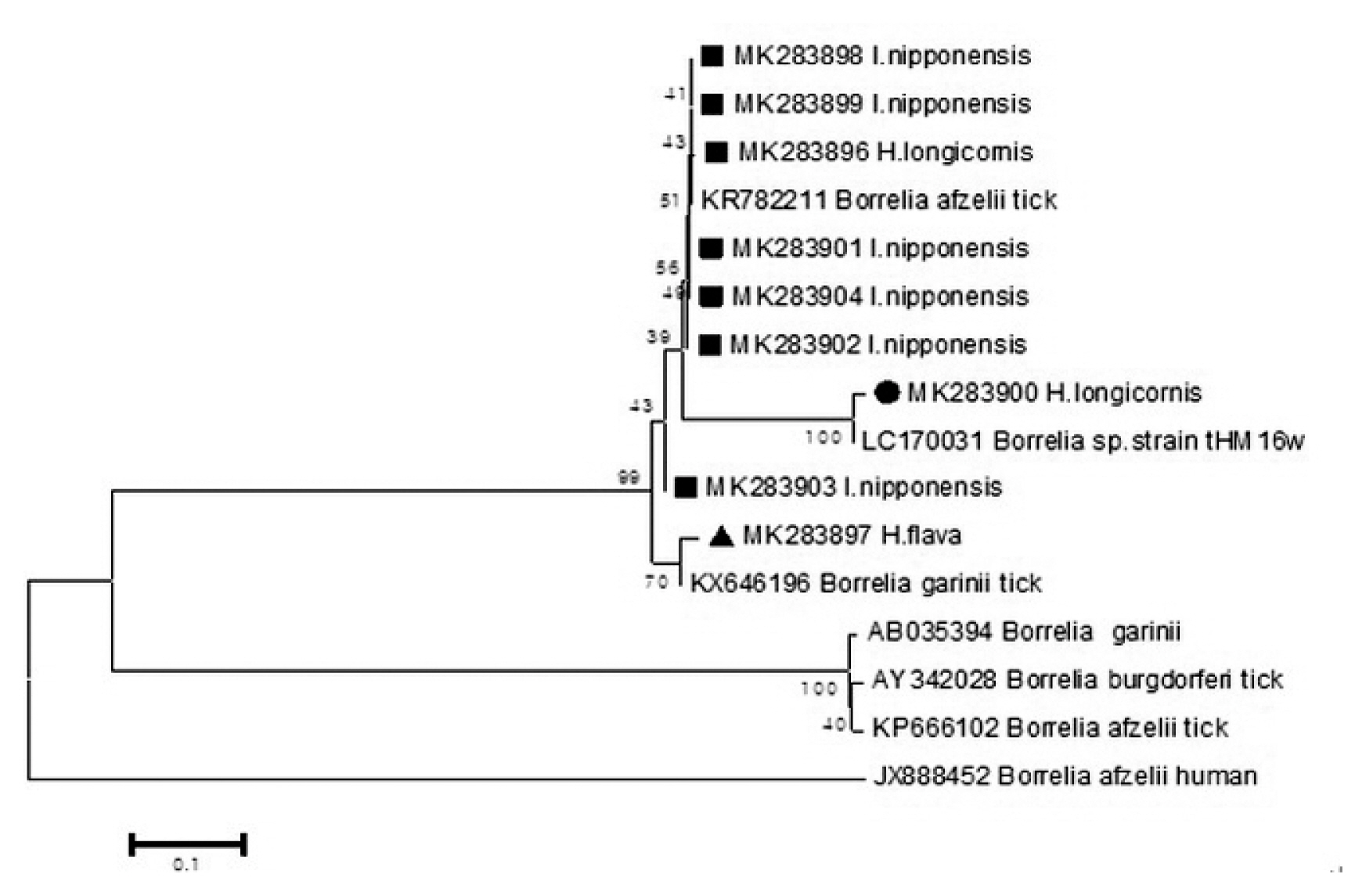
Figure & Data
References
Citations
Citations to this article as recorded by 
- Effective Methods of Estimation of Pathogen Prevalence in Pooled Ticks
Gerardo Fracasso, Marika Grillini, Laura Grassi, Francesco Gradoni, Graziana da Rold, Michela Bertola
Pathogens.2023; 12(4): 557. CrossRef - Applying next generation sequencing to detect tick-pathogens in Dermacentor nuttalli, Ixodes persulcatus, and Hyalomma asiaticum collected from Mongolia
Graham A. Matulis, Jira Sakolvaree, Bazartseren Boldbaatar, Nora Cleary, Ratree Takhampunya, B. Katherine Poole-Smith, Abigail A. Lilak, Doniddemberel Altantogtokh, Nyamdorj Tsogbadrakh, Nitima Chanarat, Nittayaphon Youngdech, Erica J. Lindroth, Jodi M. F
Ticks and Tick-borne Diseases.2023; 14(5): 102203. CrossRef - Molecular Detection of Anaplasma phagocytophilum and Ehrlichia Species in Ticks Removed from Humans in the Republic of Korea
Yu-Jung Kim, Ji Ye Seo, Seong Yoon Kim, Hee Il Lee
Microorganisms.2022; 10(6): 1224. CrossRef - Molecular Detection and Phylogeny of Tick-Borne Pathogens in Ticks Collected from Dogs in the Republic of Korea
A-Tai Truong, Jinhyeong Noh, Yeojin Park, Hyun-Ji Seo, Keun-Ho Kim, Subin Min, Jiyeon Lim, Mi-Sun Yoo, Heung-Chul Kim, Terry A. Klein, Hyunkyoung Lee, Soon-Seek Yoon, Yun Sang Cho
Pathogens.2021; 10(5): 613. CrossRef - iSeq 100 for metagenomic pathogen screening in ticks
Ju Yeong Kim, Myung-hee Yi, Alghurabi Areej Sabri Mahdi, Tai-Soon Yong
Parasites & Vectors.2021;[Epub] CrossRef - Molecular Identification of Borrelia spp. from Ticks in Pastures Nearby Livestock Farms in Korea
Haeseung Lee, Seung-Hun Lee, SungShik Shin, Dongmi Kwak
Insects.2021; 12(11): 1011. CrossRef
 PubReader
PubReader ePub Link
ePub Link Cite
Cite