
Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 8(3); 2017 > Article
-
Original Article
Hippocampus-dependent Task Improves the Cognitive Function after Ovariectomy in Rats - Songhee Cheon
-
Osong Public Health and Research Perspectives 2017;8(3):227-234.
DOI: https://doi.org/10.24171/j.phrp.2017.8.3.10
Published online: June 30, 2017
Department of Physical Therapy, College of Health Science, Youngsan University, Yangsan, Korea
- Corresponding author: Songhee Cheon, E-mail: 1000sh@ysu.ac.kr
Copyright ©2017, Korea Centers for Disease Control and Prevention
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
Abstract
-
Objectives
- Estrogen is an important hormone for cell growth, development, and differentiation by transcriptional regulation and modulation of intracellular signaling via second messengers. The reduction in the estrogen level after ovariectomy may lead to cognitive impairments associated with morphological changes in areas of the brain mediate memory. The aim of the present study was to find out the effect of tasks on the cognitive function after ovariectomy in rats.
-
Methods
- The animals used in the experiment were 50 Sprague-Dawley female rats. This study applied a hippocampus-independent task (wheel running) and a hippocampus-dependent task (Morris water maze) after ovariectomy in rats and measured the cognitive performance (object-recognition and object-location test) and growth-associated protein 43 (GAP-43) and neurotrophin 3 (NT-3) expression in the hippocampus, which is an important center for memory and learning.
-
Results
- There were meaningful differences between the hippocampus-independent and hippocampus-dependent task groups for the object-location test and GAP-43 and NT-3 expression in the hippocampus, but not the object-recognition test. However, the hippocampus-independent task group showed a significant improvement in the object-recognition test, compared to the control group.
-
Conclusion
- These results suggest that hippocampus-dependent task training after ovariectomy enhances the hippocampus-related memory and cognitive function that are associated with morphological and functional changes in the cells of the hippocampus.
- Estrogen plays a critical role in the development, maintenance, and physiology of reproductive tissues and the central nervous system (CNS) [1]. Several areas of the brain, such as the hippocampus, amygdale, and cerebral cortex, which are involved in learning and memory processes, have been shown to be extensively populated with estrogen receptors (ERs) [2]. Low levels of estrogen have been linked to increased incidence of neurodegenerative diseases and deterioration of the cognitive functions in postmenopausal women [3]. In addition, estrogen deficiency during aging may render the hippocampus more vulnerable to deterioration and exacerbate emerging age-related memory deficits. The loss of estrous cycling in middle-age has been associated with the onset of decline in the spatial memory, according to studies conducted on female rats [4] and mice [5].
- There is evidence that regular exercise has a positive effect on the memory [6]. It has been shown that short exercise programs can increase the memory and learning in young rats [7], possibly mediated by neurotrophic factors and change in the brain plasticity [8]. Exercises have effects on the hippocampus, which is an important center for spatial memory and learning [9]. Studies have shown that physical activities, including running, jumping, and walking, can increase the number of neurons, electrical activity, and neurotransmitter secretion in the hippocampus [10,11]. In addition, there are reports that hippocampus-dependent training, such as the Morris water maze training, improves the growth and survival rate of neuronal cells [12,13].
- Neurotrophic factors and transcription factors induce structural changes in the neurons of the hippocampus, which is critical for memory and learning. Neurotrophic factors such as neurotrophin 3 (NT-3) and brain-derived neurotrophic factor (BDNF), are powerful regulators of neuronal excitability and synaptic transmission, and when these factors act, synapsin I and growth-associated protein 43 (GAP-43) play important roles in neuronal and synaptic plasticity [14].
- Therefore, the principal objective of this study was to assess the effects of the types of tasks on the behavioral patterns and GAP-43 and NT-3 expression in the hippocampus, which play pivotal roles in spatial memory and learning, after ovariectomy in rats.
INTRODUCTION
- 1. Animals
- The animals used in the experiment were 50 Sprague-Dawley female rats aged 8 to 10 weeks and weighing 250 to 300 g. During the experiment, water was supplied without restriction; food was restricted, except for the given solid food and water. The vivarium temperature was maintained at 25 ± 2°C and the humidity was maintained at 60 ± 5%, and a photoperiod of 12 hours per day and a dark cycle of 12 hours were applied. The experimental animals were adapted to the environment through a two-week breeding period and subsequently subjected to experimental treatments. After ovariectomy, the animals allowed a one-week recovery period. The animals were randomly divided into 5 groups: the control (n = 10), wheel running training 1 (WT1; n = 10), wheel running training 2 (WT2; n = 10), Morris water maze training 1 (MT1; n = 10), and Morris water maze training 2 (MT2; n = 10) groups. Four groups, except the control group, were subjected to each training for 2 weeks after ovariectomy, and the WT2 and MT2 groups were also subjected to each training for 2 weeks before the operation.
- 2. Ovariectomy
- To perform ovariectomy, a general anesthetic containing a mixture of ketamine hydrochloride (Ketamine HCl; Yuhan Corp., Seoul, Korea) and Rompun (Rompun; Bayer Korea, Seoul, Korea) in the ratio of 1:1 was administered by intraperitoneal injection (0.4 mL); the abdomen was disinfected, the abdominal hair removed, and the depilated area disinfected with 10% betadine solution before the operation.
- Using surgical scissors, the abdominal muscles, skin, and peritoneum were cut and the abdominal cavity entered for approximately 2.0 cm, and the uterine tubes connected to the ovaries were ligated to the ovary with the silk thread. The ovaries were completely removed and the peritoneum, abdominal muscles, and skin were sutured. A week-long recovery period was allowed.
- 3. Wheel training
- After the ovariectomy, the animals in the wheel training group (WT1) and the front and rear wheel training group (WT2) were placed in a wheel having a diameter of 21 cm and a width of 8 cm.
- 4. Morris water maze training
- Morris water maze has been widely used as a method of assessing spatial learning and memory since its invention by Morris in 1984 [15]. In a water tank 120 cm in diameter and 50 cm in height, water was filled to a height of 40 cm and a 10 cm diameter escape platform made of a transparent acrylic plate was installed in a randomly selected quadrant. This escape platform was fixed 1 cm below the surface of the water. The water used for the maze was visibly blocked by adding milk powder to make it opaque, and the water temperature was maintained at 24°C. Morris underwater maze training group 1 (MT1) after the operation, and Morris underwater maze training group 2 (MT2) before and after the surgery were gently dropped at any point in the quadrant, and to minimize the visual information, the animals were always dropped facing the wall of the tank. At the time of the arrival, the experiment was performed for 30 seconds after ascending to the escape platform, and a time limit of 60 seconds was applied; it was performed once a day and once in each quadrant.
- 5. Object location test
- Two discriminable objects made of glass or steel among the objects used in this study, all of which were sufficiently weighted to stop a white mouse from moving, were placed 10 cm apart, along an inside edge of a box 100 cm in width, 100 cm in length and 30 cm in height for the object location test. After placing the rat in the center and allowing the rat to explore two objects for 3 minutes, the sniffing behavior within 2 cm of the object and touching the object directly with the nose or the front leg were regarded as exploring behaviors [16]. The rat was given a 5-minute break in a standard breeding field after finishing the exploration; subsequently, the rat was placed in the center of the box for 3 minutes of exploration after moving the location of one of the objects used in the prior investigation to an adjacent edge of the box. The number of times the object at an existing location and the object at the changed location were explored was measured by filming with a video camera; the results were expressed as a percentage of the exploration time of the object at a new location relative to the total exploration time, which was the sum of both the times.
- 6. Object recognition test
- The object recognition test was conducted after replacing one of the objects used in a prior exploration with a new object; all procedures were conducted in the same way as the object location test.
- 7. Western blotting
- The hippocampuses of each group were collected, washed twice in phosphate buffered saline, and subsequently homogenized and lysed with a buffer (137 mM NaCl, 8.1 mM Na2HPO4, 2.7 mM KCl, 1.5 mM KH2PO4, 2.5 mM ethylenediaminetetraacetic acid, 1 mM dithiothreitol, 0.1 mM phenylmethylsulfonyl fluoride, 10 μg/mL leupeptin [pH 7.5]) for 30minutes on ice. The lysates were centrifuged for 10 minutes at 15,000 rpm at 4°C, and the protein concentrations were determined as previously described [17]. Equal quantities of protein (40 μg) were resolved via 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. The blots were washed with TBST (10 mM TrisHCl [pH 7.6], 150 mM NaCl, 0.05% Tween 20), blocked with 5% skimmed milk for 1 hour, and subsequently incubated with primary antibodies at the dilutions recommended by the suppliers. The primary antibodies and the respective concentrations used were as follows: rabbit polyclonal antibody GAP-43 (1:3,000; Chemicon International, Temecula, CA, USA) and rabbit polyclonal antibody NT-3 (1:3,000; Santa Cruz Biotechnology, Dallas, TX, USA). The membranes were washed, and the primary antibodies were detected using horse-radish peroxidase-conjugated goat anti-rabbit immunoglobulin G (IgG) or goat-anti mouse IgG. The bands were subsequently visualized via enhanced chemiluminescence (Amersham Pharmacia Biotech, Piscataway, NJ, USA).
- 8. Statistical analysis
- The results were expressed as the means ± standard errors. All experiments were analyzed via analysis of variance (ANOVA), and comparisons of the mean values between the treatment and the control groups were analyzed via Bonferroni-Dunn’s test. The difference was regarded as statistically significant if the p-value was < 0.05.
MATERIALS AND METHODS
- 1. Object location test
- The object location test resulted in significant differences in the MT1, WT2, and MT2 groups compared to the control group (except for the WT1 group). The MT2 group showed a more significant increase than WT1 and WT2 groups (Table 1, Figure 1A).
- 2. Object recognition test
- Object recognition test resulted in more significant increases in the MT1, WT2, MT2, and WT1 groups than the control group. However, there were no significant differences among the four groups, although there were small increases (Table 1, Figure 1B).
- 3. Expression of GAP-43 and NT-3 in the hippocampus
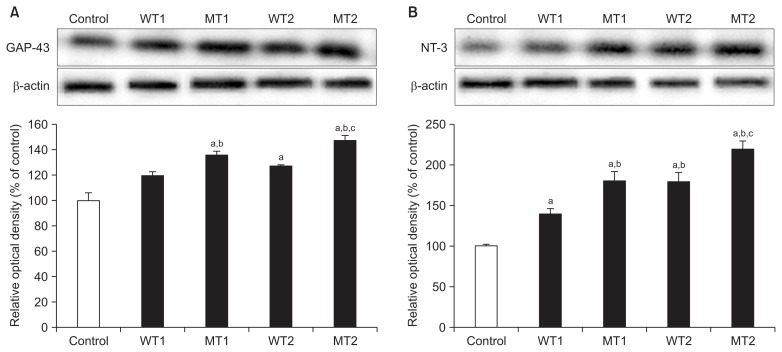
- Expression of GAP-43 in the hippocampus showed significant differences in the MT1, WT2, and MT2 groups compared to the control group, except for the WT1 group. The MT1 and MT2 groups showed significant increases compared to the WT1 group; significantly greater increase was found in the MT2 group than in the WT2 group (Figure 2A).
- The results of NT-3 expression were similar to those of GAP-43 expression. There were significant increases in the four groups, compared to the control group; the MT1, WT1, and MT2 groups showed significantly greater increases than the WT1 group (Figure 2B).
RESULTS
- This study focused on the behavioral changes related to memory and morphological changes of the hippocampus in rats whose ovaries were removed to inhibit estrogen secretion and trained with wheel running and the Morris water maze. Consequently, wheel running and Morris water maze were found to have positive effects on object recognition and object location memory. The Morris water maze seemed to particularly enhance the object-location memory. GAP-43 expression in the hippocampus increased in both WT and MT groups; however, the MT group showed a more significant increase.
- Menopause has a close relationship with memory loss and cognitive disorder. Estrogen plays an important role in neural plasticity, such as increasing CA1 dendritic spine population in the hippocampus, long-term potentiation and neurogenesis [2,18,19].
- There are numerous studies that have focused on the relationship between estrogen and memory/cognition by enhancing the brain function by estrogen injection. Xu and Zhang [20] observed the enhancing effect of long-term estradiol injection on the spatial memory of mice whose ovaries were removed, and reported the effect of estrogen on increasing the brain function. Similar to such animal studies, Sherwin [21] explained the function of estrogen in slowing down age-related cognitive loss by studying postmenopausal women.
- Monteiro et al. [22] and Feng et al. [23] showed that spatial memory was degraded in rats whose ovaries were removed; Heikkinen et al. [24] showed that long-term memory was decreased in mice whose estrogen secretion was inhibited by removing the ovary. Therefore, we also hypothesized that ovariectomy reduces memory capability and is the most commonly used method for a post-menopausal model [25].
- The hippocampus plays an important role in memory formation and learning, and this capability is reduced after menopause because of the lower estrogen level. There are numerous reports indicating that exercise facilitates the recovery of function in the CNS and increases the memory and learning ability. We applied a hippocampus-independent exercise, wheel run, to WT1 and WT2 groups. Anderson et al. [26] reported that wheel running increased the spatial learning in rats; Aiello et al. [27] reported that moderate exercise could help reduce memory-related problems when applied to postmenopausal women.
- Arida et al. [28] studied the effects of spontaneous exercise and forced exercise on the hippocampus and found that spontaneous exercise, such as wheel running, has a greater effect on enhancing the development of the hippocampus neurons. Duman et al. [29] reported that exercise not only improved the physical ability, but also acted as an antidepressant or an anxiolytic.
- We applied wheel running, a spontaneous exercise, to the WT group. Both WT1 and WT2 groups showed significant differences from the control group for the object recognition test; however, the WT1 group did not show a significant difference for the object location test.
- The MT group that was trained with the Morris water maze, on the contrary, showed a different result from the WT group. The MT group showed a greater improvement in the object recognition test, compared to the WT group; however, both the groups showed a significant difference when compared to the control group. Regarding the object location test, the MT group showed a more statistically significant improvement than the WT group.
- The main objective of the present study was to observe the effect of wheel running, a hippocampus-independent exercise, and Morris water maze, a hippocampus-dependent exercise, on the hippocampus. The hippocampus plays a critical role in various learning processes, such as spatial learning. To test such memory and learning abilities, object-recognition and object-location tests were performed, and the MT group showed a more significant improvement in both the tests. Such results indicated that an object location is related to spatial memory, which is mostly controlled by the hippocampus [30]. Also, object recognition does not rely on hippocampus, but is mostly dependent on prefrontal cortex [31], which was indicated by the result that MT group, that was trained using the Morris water maze, showed a more significant improvement in the object location test.
- Wallace et al. [32], who reported that the memory and density of dendritic spines decreased in the prefrontal cortex and hippocampus after ovariectomy, also found that the object recognition ability decreased after 1-week of ovariectomy and that the object location ability decreased after 4-weeks. Therefore, it was reported that the ovarian hormone played an important role in maintaining memory function. These results explain the reason for the greater improvement in the object location test than in the object recognition test in this study.
- Beck and Luine [33] and Bowman et al. [34] reported that ovariectomy acted as a stress, and such stress might improve the results of the object location test or other spatial memory-related tests, but does not have any effect on the object recognition tests. The fact that the WT and MT groups did not show a significant difference for the object recognition test, but showed a significant difference for the object location test indicates that hippocampus-dependent exercises have a greater effect than hippocampus-independent exercises on the object location memory.
- Wallace et al. [32] also reported that the decrease in the dendritic spine density was similar in the hippocampus and prefrontal cortex after ovariectomy. A small number of estrogen α receptors exist in the hippocampus [35] and estrogen β receptors are expressed in both the hippocampus and prefrontal cortex [36]. Therefore, it can be concluded that the interaction between estrogen and membrane receptors control the number of dendritic spines, which, in turn, is related to the memory function.
- Neural plasticity indicates the functional and physical changes of neurotransmission and the synapses. Such morphological changes of the nervous system induce changes in the signal transduction rate, which is an important factor in learning, memory, and recovery [37]. Physical exercise enhances axonal growth after a neural injury and additionally increases the expression of critical factors in the formation of axons, synapses, and axon-related peptides, such as GAP-43 [38].
- However, the effects of hippocampus-dependent and hippocampus-independent exercises on the learning and memory remain controversial. Mohapel et al. [39] indicated that hippocampus-dependent exercises had no effect or even decreased the cell survival rate when compared to hippocampus-independent exercises. On the contrary, Hairston et al. [12] reported that various types of hippocampus-dependent exercises increased the survival rates of newly-formed cells. To observe the morphological changes of the hippocampus during exercises, we quantitatively analyzed the expression of GAP-43. Subsequently, the MT group showed a more significant increase than the WT group and such a result indicated that hippocampus-dependent exercises were more effective in strengthening the hippocampus function.
- GAP-43 has been known as a direct marker for morphological changes in the dendritic spines. Gianola and Rossi [40] proved that GAP-43 was closely related to axonal sprouting and learning ability using transgenic mice models. The increase in GAP-43 expression in the MT group corresponds to the results of the hippocampus-related behavioral test.
- Holahan et al. [41] showed that GAP-43 expression controlled the spatial memory through immunohistochemistry. More particularly, the memory function improved when more GAP-43 were expressed in their phosphorylated form. Maviel et al. [42] also reported that GAP-43 controlled the process of memory formation. Pascale et al. [43] reported that Morris water maze training increased the protein and mRNA levels of GAP-43 in the hippocampus.
- Arida et al. [28] reported that wheel running and Morris water maze showed different results for neural growth and development because the exercises themselves and their protocols were different. They also indicated that physical stimulation controlled the growth process of the cells.
- Neutrophins, such as NT-3 and BDNF, play important roles in neural function, and are often used in studies related to neural plasticity. NT-3 increased following physical exercise, which also increased the synaptic plasticity. Such neurotrophic factors activate phosphate-binding proteins, such as GAP-43 that directly affect axonal growth and neurotransmission, by affecting the synaptic plasticity. However, although NT-3 plays an important role in activity-dependent neural plasticity, control of expression through neuromuscular activity is very complex when compared to other neurotrophic factors; moreover, it also depends on the activity load [14].
- Similar to the results of GAP-43, NT-3 expression also increased more significantly in the MT group than in the WT group. However, the WT group also showed a significant increase in expression when compared to the control group. Such results indicated that wheel running and Morris water maze provided sufficient afferent input to both WT and MT groups.
- Continuous application of hippocampus-dependent and hippocampus-independent exercises after ovariectomy increased the hippocampus-related performance level. However, hippocampus-dependent exercises seemed to have a greater effect on the object location test than on the object recognition test. This corresponds to the histological changes in the hippocampus that were observed via GAP-43 expression. Therefore, depending on the form of exercise and tasks, changes in the cognitive function related to the hippocampus can be observed, and such functional changes are also related to physiological changes in the neurons.
DISCUSSION
-
Acknowledgements
- This research was supported by a Youngsan University Research Grant in 2017.
ACKNOWLEDGMENTS
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
Article information
- 1. Gruber CJ, Tschugguel W, Schneeberger C, et al. Production and actions of estrogens. N Engl J Med 2002;346:340−52. https://doi.org/10.1056/NEJMra000471. PMID: 10.1056/NEJMra000471. PMID: 11821512.ArticlePubMed
- 2. Shughrue PJ, Lane MV, Merchenthaler I. Comparative distribution of estrogen receptor-alpha and -beta mRNA in the rat central nervous system. J Comp Neurol 1997;388:507−25. https://doi.org/10.1002/(SICI)1096-9861(19971201)388:4<507::AID-CNE1>3.0.CO;2-6. PMID: 10.1002/(SICI)1096-9861(19971201)388:4<507::AID-CNE1>3.0.CO;2-6. PMID: 9388012.ArticlePubMed
- 3. Kawas C, Resnick S, Morrison A, et al. A prospective study of estrogen replacement therapy and the risk of developing Alzheimer’s disease: the Baltimore Longitudinal Study of Aging. Neurology 1997;48:1517−21. https://doi.org/10.1097/00006254-199711000-00020. PMID: 10.1212/WNL.48.6.1517. PMID: 9191758.ArticlePubMed
- 4. Markowska AL. Sex dimorphisms in the rate of age-related decline in spatial memory: relevance to alterations in the estrous cycle. J Neurosci 1999;19:8122−33. PMID: 10479712.ArticlePubMedPMC
- 5. Frick KM, Burlingame LA, Arters JA, et al. Reference memory, anxiety and estrous cyclicity in C57BL/6NIA mice are affected by age and sex. Neuroscience 2000;95:293−307. https://doi.org/10.1016/S0306-4522(99)00418-2. PMID: 10.1016/S0306-4522(99)00418-2. PMID: 10619486.ArticlePubMed
- 6. Cotman CW, Berchtold NC. Exercise: a behavioral intervention to enhance brain health and plasticity. Trends Neurosci 2002;25:295−301. https://doi.org/10.1016/S0166-2236(02)02143-4. PMID: 10.1016/S0166-2236(02)02143-4. PMID: 12086747.ArticlePubMed
- 7. Uysal N, Tugyan K, Kayatekin BM, et al. The effects of regular aerobic exercise in adolescent period on hippocampal neuron density, apoptosis and spatial memory. Neurosci Lett 2005;383:241−5. https://doi.org/10.1016/j.neulet.2005.04.054. PMID: 10.1016/j.neulet.2005.04.054. PMID: 15878799.ArticlePubMed
- 8. Neeper SA, Gómez-Pinilla F, Choi J, et al. Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain Res 1996;726:49−56. https://doi.org/10.1016/0006-8993(96)00273-9. PMID: 10.1016/0006-8993(96)00273-9. PMID: 8836544.ArticlePubMed
- 9. Tong L, Shen H, Perreau VM, et al. Effects of exercise on gene-expression profile in the rat hippocampus. Neurobiol Dis 2001;8:1046−56. https://doi.org/10.1006/nbdi.2001.0427. PMID: 10.1006/nbdi.2001.0427. PMID: 11741400.ArticlePubMed
- 10. Kim YP, Kim H, Shin MS, et al. Age-dependence of the effect of treadmill exercise on cell proliferation in the dentate gyrus of rats. Neurosci Lett 2004;355:152−4. https://doi.org/10.1016/j.neulet.2003.11.005. PMID: 10.1016/j.neulet.2003.11.005. PMID: 14729257.ArticlePubMed
- 11. Ahmadiasl N, Alaei H, Hänninen O. Effect of exercise on learning, memory and levels of epinephrine in rats’ hippocampus. J Sports Sci Med 2003;2:106−9. PMID: 24627662.PubMedPMC
- 12. Hairston IS, Little MT, Scanlon MD, et al. Sleep restriction suppresses neurogenesis induced by hippocampus-dependent learning. J Neurophysiol 2005;94:4224−33. https://doi.org/10.1152/jn.00218.2005. PMID: 10.1152/jn.00218.2005. PMID: 16014798.ArticlePubMed
- 13. Olariu A, Cleaver KM, Shore LE, et al. A natural form of learning can increase and decrease the survival of new neurons in the dentate gyrus. Hippocampus 2005;15:750−62. https://doi.org/10.1002/hipo.20097. PMID: 10.1002/hipo.20097. PMID: 16010660.ArticlePubMed
- 14. Gómez-Pinilla F, Ying Z, Roy RR, et al. Afferent input modulates neurotrophins and synaptic plasticity in the spinal cord. J Neurophysiol 2004;92:3423−32. https://doi.org/10.1152/jn.00432.2004. PMID: 10.1152/jn.00432.2004. PMID: 15548637.ArticlePubMed
- 15. Silhol M, Arancibia S, Maurice T, et al. Spatial memory training modifies the expression of brain-derived neurotrophic factor tyrosine kinase receptors in young and aged rats. Neuroscience 2007;146:962−73. https://doi.org/10.1016/j.neuroscience.2007.02.013. PMID: 10.1016/j.neuroscience.2007.02.013. PMID: 17391859.ArticlePubMed
- 16. Bisagno V, Ferguson D, Luine VN. Chronic D-amphetamine induces sexually dimorphic effects on locomotion, recognition memory, and brain monoamines. Pharmacol Biochem Behav 2003;74:859−67. https://doi.org/10.1016/S0091-3057(03)00017-0. PMID: 10.1016/S0091-3057(03)00017-0. PMID: 12667900.ArticlePubMed
- 17. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976;72:248−54. https://doi.org/10.1016/0003-2697(76)90527-3. PMID: 10.1016/0003-2697(76)90527-3. PMID: 942051.ArticlePubMed
- 18. Shilling V, Jenkins V, Fallowfield L, et al. The effects of oestrogens and anti-oestrogens on cognition. Breast 2001;10:484−91. https://doi.org/10.1054/brst.2001.0311. PMID: 10.1054/brst.2001.0311. PMID: 14965628.ArticlePubMed
- 19. Maki PM. Hormone therapy and cognitive function: is there a critical period for benefit? Neuroscience 2006;138:1027−30. https://doi.org/10.1016/j.neuroscience.2006.01.001. PMID: 10.1016/j.neuroscience.2006.01.001. PMID: 16488547.ArticlePubMed
- 20. Xu X, Zhang Z. Effects of estradiol benzoate on learning-memory behavior and synaptic structure in ovariectomized mice. Life Sci 2006;79:1553−60. https://doi.org/10.1016/j.lfs.2006.04.020. PMID: 10.1016/j.lfs.2006.04.020. PMID: 16750837.ArticlePubMed
- 21. Sherwin BB. Estrogen and cognitive functioning in women. Endocr Rev 2003;24:133−51. https://doi.org/10.1210/er.2001-0016. PMID: 10.1210/er.2001-0016. PMID: 12700177.ArticlePubMed
- 22. Monteiro SC, Matté C, Bavaresco CS, et al. Vitamins E and C pre-treatment prevents ovariectomy-induced memory deficits in water maze. Neurobiol Learn Mem 2005;84:192−9. https://doi.org/10.1016/j.nlm.2005.08.002. PMID: 10.1016/j.nlm.2005.08.002. PMID: 16169259.ArticlePubMed
- 23. Feng Z, Cheng Y, Zhang JT. Long-term effects of melatonin or 17 beta-estradiol on improving spatial memory performance in cognitively impaired, ovariectomized adult rats. J Pineal Res 2004;37:198−206. https://doi.org/10.1111/j.1600-079X.2004.00158.x. PMID: 10.1111/j.1600-079X.2004.00158.x. PMID: 15357665.ArticlePubMed
- 24. Heikkinen T, Puoliväli J, Tanila H. Effects of long-term ovariectomy and estrogen treatment on maze learning in aged mice. Exp Gerontol 2004;39:1277−83. https://doi.org/10.1016/j.exger.2004.05.005. PMID: 10.1016/j.exger.2004.05.005. PMID: 15489050.ArticlePubMed
- 25. Savonenko AV, Markowska AL. The cognitive effects of ovariectomy and estrogen replacement are modulated by aging. Neuroscience 2003;119:821−30. PMID: 10.1016/S0306-4522(03)00213-6. PMID: 12809703.ArticlePubMed
- 26. Anderson BJ, Rapp DN, Baek DH, et al. Exercise influences spatial learning in the radial arm maze. Physiol Behav 2000;70:425−9. https://doi.org/10.1016/S0031-9384(00)00282-1. PMID: 10.1016/S0031-9384(00)00282-1. PMID: 11110995.ArticlePubMed
- 27. Aiello EJ, Yasui Y, Tworoger SS, et al. Effect of a yearlong, moderate-intensity exercise intervention on the occurrence and severity of menopause symptoms in postmenopausal women. Menopause 2004;11:382−8. https://doi.org/10.1097/01.GME.0000113932.56832.27. PMID: 10.1097/01.GME.0000113932.56832.27. PMID: 15243275.ArticlePubMed
- 28. Arida RM, Scorza CA, da Silva AV, et al. Differential effects of spontaneous versus forced exercise in rats on the staining of parvalbumin-positive neurons in the hippocampal formation. Neurosci Lett 2004;364:135−8. https://doi.org/10.1016/j.neulet.2004.03.086. PMID: 10.1016/j.neulet.2004.03.086. PMID: 15196661.ArticlePubMed
- 29. Duman CH, Schlesinger L, Russell DS, et al. Voluntary exercise produces antidepressant and anxiolytic behavioral effects in mice. Brain Res 2008;1199:148−58. https://doi.org/10.1016/j.brainres.2007.12.047. PMID: 10.1016/j.brainres.2007.12.047. PMID: 18267317.ArticlePubMedPMC
- 30. Broadbent NJ, Squire LR, Clark RE. Spatial memory, recognition memory, and the hippocampus. Proc Natl Acad Sci U S A 2004;101:14515−20. https://doi.org/10.1073/pnas.0406344101. PMID: 10.1073/pnas.0406344101. PMID: 15452348.ArticlePubMedPMC
- 31. Ennaceur A, Neave N, Aggleton JP. Spontaneous object recognition and object location memory in rats: the effects of lesions in the cingulate cortices, the medial prefrontal cortex, the cingulum bundle and the fornix. Exp Brain Res 1997;113:509−19. PMID: 10.1007/PL00005603. PMID: 9108217.ArticlePubMed
- 32. Wallace M, Luine V, Arellanos A, et al. Ovariectomized rats show decreased recognition memory and spine density in the hippocampus and prefrontal cortex. Brain Res 2006;1126:176−82. https://doi.org/10.1016/j.brainres.2006.07.064. PMID: 10.1016/j.brainres.2006.07.064. PMID: 16934233.ArticlePubMed
- 33. Beck KD, Luine VN. Sex differences in behavioral and neuro-chemical profiles after chronic stress: role of housing conditions. Physiol Behav 2002;75:661−73. https://doi.org/10.1016/S0031-9384(02)00670-4. PMID: 10.1016/S0031-9384(02)00670-4. PMID: 12020731.ArticlePubMed
- 34. Bowman RE, Beck KD, Luine VN. Chronic stress effects on memory: sex differences in performance and monoaminergic activity. Horm Behav 2003;43:48−59. https://doi.org/10.1016/S0018-506X(02)00022-3. PMID: 10.1016/S0018-506X(02)00022-3. PMID: 12614634.ArticlePubMed
- 35. Zhang JQ, Cai WQ, Zhou DS, et al. Distribution and differences of estrogen receptor beta immunoreactivity in the brain of adult male and female rats. Brain Res 2002;935:73−80. https://doi.org/10.1016/S0006-8993(02)02460-5. PMID: 10.1016/S0006-8993(02)02460-5. PMID: 12062475.ArticlePubMed
- 36. Kritzer MF. Regional, laminar and cellular distribution of immunoreactivity for ERbeta in the cerebral cortex of hormonally intact, postnatally developing male and female rats. Cereb Cortex 2006;16:1181−92. https://doi.org/10.1093/cercor/bhj059. PMID: 10.1093/cercor/bhj059. PMID: 16221921.ArticlePubMed
- 37. Frey D, Laux T, Xu L, et al. Shared and unique roles of CAP23 and GAP43 in actin regulation, neurite outgrowth, and anatomical plasticity. J Cell Biol 2000;149:1443−54. https://doi.org/10.1083/jcb.149.7.1443. PMID: 10.1083/jcb.149.7.1443. PMID: 10871284.ArticlePubMedPMC
- 38. Vaynman S, Ying Z, Gómez-Pinilla F. Exercise induces BDNF and synapsin I to specific hippocampal subfields. J Neurosci Res 2004;76:356−62. https://doi.org/10.1002/jnr.20077. PMID: 10.1002/jnr.20077. PMID: 15079864.ArticlePubMed
- 39. Mohapel P, Mundt-Petersen K, Brundin P, et al. Working memory training decreases hippocampal neurogenesis. Neuroscience 2006;142:609−13. https://doi.org/10.1016/j.neuroscience.2006.07.033. PMID: 10.1016/j.neuroscience.2006.07.033. PMID: 16962715.ArticlePubMed
- 40. Gianola S, Rossi F. GAP-43 overexpression in adult mouse Purkinje cells overrides myelin-derived inhibition of neurite growth. Eur J Neurosci 2004;19:819−30. https://doi.org/10.1111/j.0953-816X.2004.03190.x. PMID: 10.1111/j.0953-816X.2004.03190.x. PMID: 15009129.ArticlePubMed
- 41. Holahan MR, Honegger KS, Tabatadze N, et al. GAP-43 gene expression regulates information storage. Learn Mem 2007;14:407−15. https://doi.org/10.1101/lm.581907. PMID: 10.1101/lm.581907. PMID: 17554085.ArticlePubMedPMC
- 42. Maviel T, Durkin TP, Menzaghi F, et al. Sites of neocortical reorganization critical for remote spatial memory. Science 2004;305:96−9. https://doi.org/10.1126/science.1098180. PMID: 10.1126/science.1098180. PMID: 15232109.ArticlePubMed
- 43. Pascale A, Gusev PA, Amadio M, et al. Increase of the RNA-binding protein HuD and posttranscriptional up-regulation of the GAP-43 gene during spatial memory. Proc Natl Acad Sci U S A 2004;101:1217−22. https://doi.org/10.1073/pnas.0307674100. PMID: 10.1073/pnas.0307674100. PMID: 14745023.ArticlePubMedPMC
REFERENCES
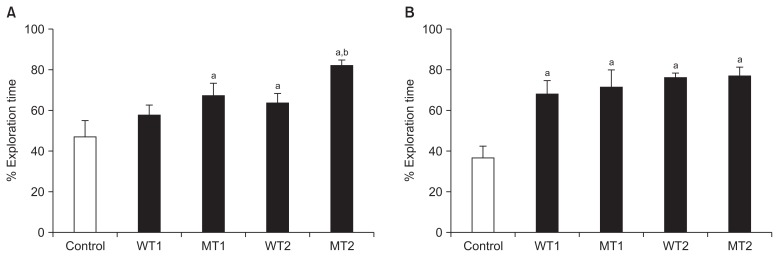
| Control | WT1 | MT1 | WT2 | MT2 | |
|---|---|---|---|---|---|
| Location | 46.85 ± 7.99a | 57.59 ± 4.96ab | 67.03 ± 6.28bc | 63.56 ± 4.56b | 81.92 ± 2.80c |
| Recognition | 36.34 ± 5.94a | 67.90 ± 6.79b | 71.24 ± 8.32b | 75.73 ± 2.59b | 76.79 ± 4.43b |
Values are presented as mean ± standard error.
WT1, group applied wheel running exercise for 2 weeks after ovariectomy; WT2, wheel running exercise for 2 weeks before ovariectomy and 2 weeks after ovariectomy; MT1, Morris water maze training for 2 weeks after ovariectomy; MT2, Morris water maze training for 2 weeks before ovariectomy and 2 weeks after ovariectomy.
abc Values with different superscripts in the same column are different significantly (p < 0.05) by Duncan’s multiple range test.
Figure & Data
References
Citations
- Radiomic features of the hippocampal based on magnetic resonance imaging in the menopausal mouse model linked to neuronal damage and cognitive deficits
Jie Zhao, Yan Jiao, Hui Wang, Peiji Song, Zhen Gao, Xue Bing, Chunling Zhang, Aimei Ouyang, Jian Yao, Song Wang, Huijie Jiang
Brain Imaging and Behavior.2023;[Epub] CrossRef - Targeting the TLR4/NF-κΒ Axis and NLRP1/3 Inflammasomes by Rosuvastatin: A Role in Impeding Ovariectomy-Induced Cognitive Decline Neuropathology in Rats
Muhammed A. Saad, Muhammad Y. Al-Shorbagy, Hany H. Arab
Molecular Neurobiology.2022; 59(7): 4562. CrossRef -
Estrogen Exerts Neuroprotective Effects in Vascular Dementia Rats by Suppressing Autophagy and Activating the Wnt/β-Catenin Signaling Pathway
Yanyan Yang, Lei Zhao, Na Li, Congwei Dai, Nan Yin, Zhaoping Chu, Xiaoyan Duan, Xiaoli Niu, Ping Yan, Peiyuan Lv
Neurochemical Research.2020; 45(9): 2100. CrossRef - Modulation of aromatase by natural compounds—A pharmacophore guided molecular modelling simulations
S. Rampogu, C. Park, M. Son, A. Baek, A. Zeb, G. Lee, K.W. Lee
South African Journal of Botany.2019; 120: 230. CrossRef - Early Stage Glycosylation Biomarkers in Alzheimer’s Disease
Patricia Regan, Paula L. McClean, Thomas Smyth, Margaret Doherty
Medicines.2019; 6(3): 92. CrossRef
 PubReader
PubReader Cite
Cite

