
Articles
- Page Path
- HOME > Osong Public Health Res Perspect > Volume 8(1); 2017 > Article
-
Original Article
The Effect ofLactobacillus acidophilus PTCC 1643 on Cultured Intestinal Epithelial Cells Infected withSalmonella enterica serovar Enteritidis - Mona Moshiria, Mohammad Mehdi Soltan Dallala,b, Farhad Rezaeic, Masoumeh Douraghia, Laleh Sharifid, Zahra Noroozbabaeic, Mehrdad Gholamie, Abbas Mirshafieyf
-
Osong Public Health and Research Perspectives 2017;8(1):54-60.
DOI: https://doi.org/10.24171/j.phrp.2017.8.1.07
Published online: February 28, 2017
aDivision of Microbiology, Department of Pathobiology, School of Public Health, Tehran University of Medical Sciences, Tehran, Iran
bFood Microbiology Research Center, Tehran University of Medical Sciences, Tehran, Iran
cDepartment of Virology, Tehran University of Medical Sciences, Tehran, Iran
dResearch Center for Immunodeficiencies, Tehran University of Medical Sciences, Tehran, Iran
eDepartment of Microbiology, Faculty of Medicine, Iran University of Medical Sciences, Tehran, Iran
fDepartment of Immunology, School of Public Health, Tehran University of Medical Sciences, Tehran, Iran
- Corresponding author: Mohammad Mehdi Soltan Dallal, E-mail: msoltandallal@gmail.com
Copyright © 2017 Korea Centers for Disease Control and Prevention
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
Abstract
-
Objectives
- Gastrointestinal disorders caused by Salmonella enterica serovar Enteritidis (SesE) are a significant health problem around the globe. Probiotic bacteria have been shown to have positive effects on the immune responses. Lactobacillus acidophilus was examined for its capability to influence the innate immune response of HT29 intestinal epithelial cells towards SesE. The purpose of this work was to assess the effect of L. acidophilus PTCC 1643 on cultured intestinal epithelial cells infected with SesE.
-
Methods
- HT29 cells were cultured in Roswell Park Memorial Institute medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. The cells were treated with L. acidophilus PTCC 1643 after or before challenge with SesE. At 2 and 4 hours post-infection, we measured changes in the expression levels of TLR2 and TLR4 via real-time polymerase chain reaction.
-
Results
- Treatment with L. acidophilus inhibited SesE-induced increases in TLR2 and TLR4 expression in the infected HT29 cells. Moreover, the expression of TLR2 and TLR4 in cells that were pretreated with L. acidophilus and then infected with SesE was significantly higher than that in cells infected with SesE without pretreatment. Taken together, the results indicated that L. acidophilus had an anti-inflammatory effect and modulated the innate immune response to SesE by influencing TLR2 and TLR4 expression.
-
Conclusion
- Our findings suggested that L. acidophilus PTCC 1643 was able to suppress inflammation caused by SesE infection in HT29 cells and reduce TLR2 and TLR4 expression. Additional in vivo and in vitro studies are required to further elucidate the mechanisms underlying this anti-inflammatory effect.
- Probiotic strains are live bacteria and yeasts that have beneficial effects on human health when ingested, particularly for the digestive system [1]. They are capable of changing the population of flora in the gut and preventing intestinal infections, gut cancer, and allergy, and they have specific immunomodulatory properties in the gut that are mediated by the repression of inflammatory mediators. Additionally, they can influence the properties of the intestinal barrier [2,3]. The functional effects of probiotics are correlated with their capabilities to compete with pathogenic organisms for adhesion positions and adjust the host’s immune response [4]. Among the many health benefits attributed to probiotics, their beneficial interactions with the immune system are supported by a growing corpus of in vivo and in vitro experiments as well as clinical evidence [5]. Lactic acid bacteria, such as lactobacilli and bifidobacteria, are usual residents of the human gastrointestinal tract and are commonly used in dairy products [6]. Salmonella is known as a major causative agent of food-borne illness in developed and developing countries [7]. Salmonella enterica serovar Enteritidis (SesE) is the predominant cause of food-borne salmonellosis in many countries including Iran [8]. Previous studies demonstrated that Lactobacillus spp. have immunomodulatory effects and antagonistic effects against Salmonella infection both in vitro and in vivo [9–11]. Many of the effects of probiotic strains are mediated via immune regulation, especially through modulating the balance between pro- and anti-inflammatory cytokines [12]. Host pattern recognition receptors, such as Toll-like receptors (TLRs), play a key role in the recognition of pathogen components and the response to specific pathogen-associated molecular patterns that are associated with diverse microorganisms including bacteria, viruses, and fungi [13]. Hence, the induction of TLR2 and TLR4 after infection with intestinal bacteria may lead to the overexpression of inflammatory cytokines through the activation of their corresponding signaling pathways [14]. Lactobacillus strains are able to suppress the activation of TLR2 and TLR4 signaling. Furthermore, some probiotic strains have been shown to increase phagocytosis in phagocytic cells and modify cytokine production following interactions with TLRs in different cell populations, although the studies are limited to date and the results have sometimes been contradictory [15]. Thus, the purpose of this work was to assess the effect of the probiotic L. acidophilus PTCC 1643 on cultured intestinal epithelial cells infected with SesE in vitro.
INTRODUCTION
- 1. Bacterial preparation
- This study was conducted in the Microbiology Laboratory of the Department of Microbiology of Tehran University of Medical Sciences in Tehran, Iran, during 2014 and 2015. L. acidophilus PTCC 1643 was purchased from the Persian Type Culture Collection of the Iranian Research Organization for Science and Technology in Tehran, Iran. L. acidophilus was cultured in de Man, Rogosa, and Sharpe broth (Merck, Darmstadt, Germany) for 48 hours under a 5% CO2 atmosphere at 37°C. Stock cultures were maintained at −20°C in 15% glycerol. SesE ATCC 17036 was cultured in Luria–Bertani medium (Merck) at 37°C overnight [16].
- 2. Cell culture
- In this study, the HT29 human colon adenocarcinoma cell line was purchased from the Cell Bank at the International Genetic and Biological Center (Tehran, Iran), and grown in 25 cm2 cell culture flasks (Greiner Bio-one, Frickenhausen, Germany) in an incubator with 95% (v/v) humidified air containing 5% CO2 at 37°C for 4–5 days. The cells were cultured in Roswell Park Memorial Institute 1640 medium (RPMI 1640; HyClone, Logan, UT, USA) supplemented with 10% (v/v) inactivated fetal bovine serum and 1% (v/v) penicillin/streptomycin (Invitrogen, Carlsbad, CA, USA). A negative control (untreated HT29 cells) was used in all experiments.
- 3. Co-culture of L. acidophilus and SesE with HT29 cells
- HT29 cells were seeded into 12-well plates at a density of 1 × 106 cells/well. After 2–3 hours of incubation, the culture medium was replaced with fresh RPMI 1640 medium (without penicillin/streptomycin). HT29 cells were co-incubated with L. acidophilus at a density of 1 × 107 colony-forming units (CFU)/well for 2 hours at 37°C under a 5% CO2 atmosphere. In separate 12-well plates, HT29 cells were infected with SesE at a density of 1 × 107 CFU/well for 2 hours at 37°C under a 5% CO2 atmosphere. After incubation, the cells were washed twice with culture media (RPMI 1640 without penicillin/streptomycin and fetal calf serum) and 150 μL of SesE (1 × 107 CFU/well) was added to a plate containing HT29 and L. acidophilus, then 150 μL of L. acidophilus (1 × 107 CFU/well) was added to a plate containing HT29 and SesE. Plates incubated with either L. acidophilus or SesE alone were also included in the experiment. In the final step, the plates were rotated gently for 10 seconds and then incubated at 37°C for 2–4 hours. After incubation, HT29 cells were collected via 5 minutes centrifugation at 5,200 × g at room temperature. Each cell pellet was washed twice with sterile phosphate-buffered saline (pH 6.7) to quantify the expression levels of TLR2 and TLR4.
- 4. RNA extraction and cDNA synthesis
- Total RNAs were extracted from HT29 cells using the QIAzol® Lysis Reagent (Qiagen, Hilden, Germany). The quantity and purity of the RNA were assessed by measuring the absorbance at 260 nm and its ratio relative to that at 280 nm using an ultraviolet spectrophotometer (PhotoBiometer; Eppendorf, Hamburg, Germany). In the next step, total RNA was treated with RNase-free DNase I (Thermo Fisher Scientific, Waltham, MA, USA) to eliminate DNA contaminants and its integrity was checked by electrophoresis on an agarose gel stained with GelRed™ (Biotium, Hayward, CA, USA). For real-time (RT) polymerase chain reaction (PCR) experiments, cDNA was synthesized from total RNA using the QuantiTect® Reverse Transcription kit (Qiagen) according to the manufacturer’s recommendations. The primer sequences used for this study are shown in Table 1. The β-actin gene (ACTB) was used as a housekeeping reference gene.
- 5. Quantitative RT-PCR
- To characterize the effects of L. acidophilus on the expression of TLR2 and TLR4, RT-PCR was carried out using SYBR® Premix Ex Taq™ (Takara Biotechnology, Otsu, Japan) on a thermal cycler (StepOnePlus™; Applied Biosystems, Foster City, CA, USA) during 40 cycles. Briefly, 20 μL of reaction mixture contained 5 μL cDNA, 10 μL Power SYBR® Green PCR master mix (Applied Biosystems), 4 μl RNase-free water, and 0.5 μL each of forward and reverse primers. The PCRs were performed with 1 cycle at 95°C for 10 minutes for initial denaturation followed by 40 cycles at 95°C for 30 seconds and 60°C for 30 seconds, then a final extension step for 30 seconds at 72°C. The results were expressed as mean values averaged from three independent experiments. ACTB was used as an endogenous control. In all tests, a negative control was used to determine contamination. The relative quantity (RQ) of gene expression for the sample was calculated using the 2−ΔΔCt method.
- 6. Statistical analysis
- Statistical analyses were performed using the Microsoft Excel 2010 (Microsoft, Redmond, WA, USA) analysis tool package. Unless otherwise stated, all results are the mean values ± standard deviation of two independent experiments with at least 3 replicates. Data from each experiment were analyzed by one-way analysis of variance. Differences at p < 0.05 were considered to be statistically significant. Differences at p < 0.01 were considered to be statistically significant.
- 7. Ethics statement
- Ethical approval was not required for the study, as there was no direct patient involvement and only cell cultures and bacterial strains were studied.
MATERIALS AND METHODS
- 1. Effects of L. acidophilus and SesE on TLR2 expression in HT29 cells
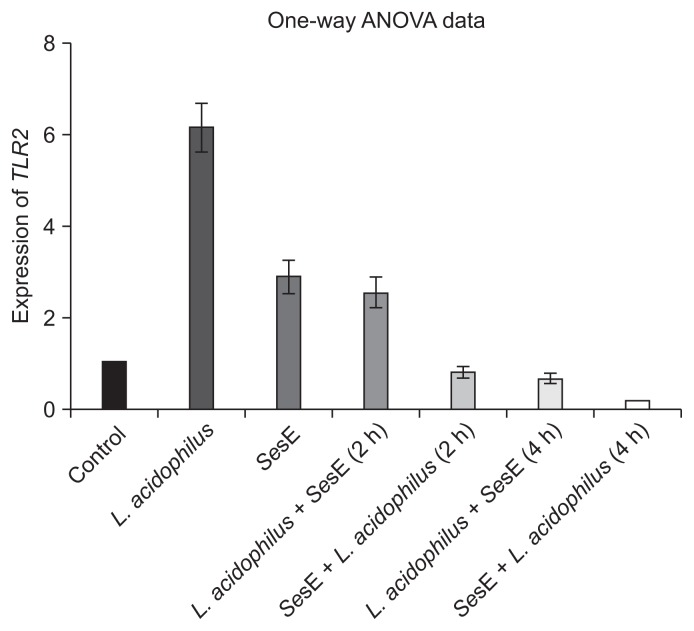
- Analysis by RT-PCR revealed that the RQ of TLR2 in HT29 cells after 6 hours of incubation with SesE was 2.89, while that in the cells treated with L. acidophilus (without SesE) was 6.13 (p < 0.01). Our results indicated that TLR2 expression in HT29 cells exposed to L. acidophilus or SesE was elevated compared with that of the control, and the increase in cells treated with L. acidophilus was higher than that in the cells infected with SesE (Figure 1).
- 2. Effects of co-culture with L. acidophilus and SesE on TLR2 expression in HT29 cells
- When HT29 cells were incubated with SesE for 2 hours and then treated with L. acidophilus, the RQ of TLR2 expression after 2 hours treatment with L. acidophilus was 0.785, and after 4 hours treatment, the RQ decreased to 0.146. When the HT29 cells were treated with L. acidophilus and then incubated with SesE, the RQs of TLR2 expression after 2 and 4 hours incubation with SesE were 2.55 and 0.650, respectively. The most notable of these findings was that TLR2 expression in SesE-exposed HT29 cells was markedly reduced after 4 hours of incubation with L. acidophilus (Figure 2).
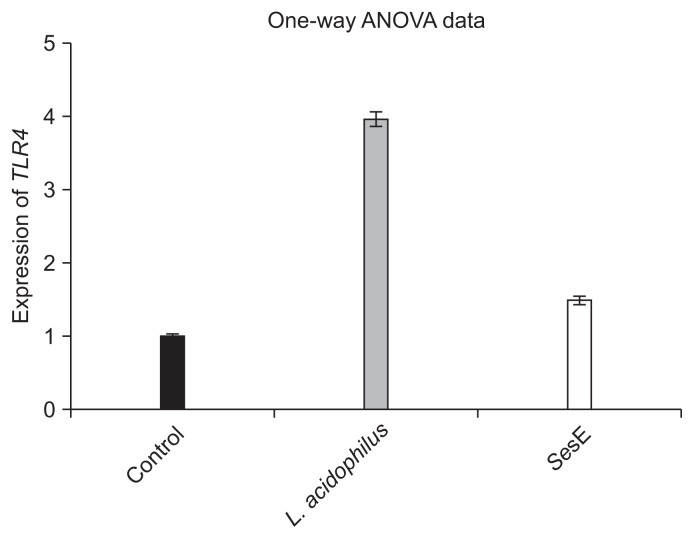
- 3. Effects of L. acidophilus and SesE on TLR4 expression in HT29 cells
- TLR4 expression was investigated in HT29 cells exposed to either L. acidophilus or SesE. The RQ of TLR4 expression in the HT29 cells infected with SesE was 1.489 after 6 hours incubation, while that in the cells treated with L. acidophilus was 3.97 (p < 0.001). These findings indicated that L. acidophilus increased the expression of TLR4 in the HT29 cells to a greater extent than SesE did (Figure 3).
- 4. Effects of co-culture with L. acidophilus and SesE on TLR4 expression in HT29 cells
- To further examine the effects of L. acidophilus and SesE on TLR4 expression in HT29 cells, RT-PCR was performed. When HT29 cells were incubated with SesE for 2 hours and then treated with L. acidophilus, the RQs of TLR4 expression at 2 and 4 hours after treatment with L. acidophilus were 0.623 and 0.414, respectively.
- When HT29 cells were treated with L. acidophilus for 2 hours and then incubated with SesE, TLR4 expression significantly increased after 2 hours exposure to SesE (RQ = 1.432) but was markedly reduced after 4 hours exposure to SesE (RQ = 0.840). Taken together, the results showed that the expression levels of TLR2 and TLR4 were reduced after treatment with L. acidophilus in HT29 cells pre-incubated with SesE, and TLR4 was downregulated more than TLR2 (Figure 4).
RESULTS
- Food-borne diseases caused by SesE remain a major public health concern. SesE is one of the predominant bacteria that cause human salmonellosis in many developing countries including Iran [8]. Recently, probiotics have been effectively used for the treatment and inhibition of enteric infections in humans [17]. Among the normal human gastrointestinal flora, L. acidophilus plays a key role in exerting inhibitory effects on the growth of obligate pathogens such as SesE, Staphylococcus aureus, Shigella dysenteriae, and other microorganisms [18,19]. Hence, previous studies have shown that L. acidophilus possesses a strong anti-inflammatory activity [20,21].
- Previous studies have reported conflicting results about the expression of TLR2 in intestinal epithelial cells. Melmed et al [22] showed that intestinal epithelial cells were largely inattentive to bacteria that are recognized via TLR2. Otte et al [23] reported that intestinal epithelial cell lines showed reduced TLR4 and TLR2 expression during prolonged contact with TLR ligands (lipopolysaccharides and lipoteichoic acid). Furrie et al [24] reported that the expression of TLR2 and TLR4 was significantly increased in HT29 cells when they were cocultured with certain gram-positive bacteria, but gram-negative bacteria elicited no such change in expression. TLR2 stimulation can protect the epithelial barrier, and it has been shown that the activation of TLR2 plays an essential role in mediating resistance to bacterial invasion by enhancing the innate immune response [13]. Vizoso Pinto et al [25] reported that TLR9 and TLR2 were expressed by HT29 cells and their expression ratio changed after exposure to Lactobacillus plantarum BFE 1685, but was unchanged after incubation with S. enterica serovar Typhimurium only. Similar results were also obtained by Bermudez-Brito and coworkers [26]. In addition, other investigations have suggested that probiotics alone can promote the expression of TLR2 and activate the innate immune response and immunostimulation [26–28].
- In our study, the expression of TLR2 significantly decreased in HT29 cells when they were treated with L. acidophilus either after or before infection with SesE. Our observation that L. acidophilus modulates TLR gene expression is in agreement with several studies conducted in Germany, Spain, and France [25,26,29].
- Our results revealed that TLR4 expression in HT29 cells increased after treatment with L. acidophilus alone. Our findings agree with those previously reported by a study performed in Argentina [28]. In contrast, previous studies have shown that TLR4 expression in Caco-2 cells decreased after treatment with lactic acid bacteria [30,31], whereas Miettinen et al [32] found that Lactobacillus rhamnosus GG did not influence TLR4 expression.
- In the present study, the expression of TLR4 was decreased in HT29 cells when they were treated with L. rhamnosus either before or after infection with SesE. The expression of both TLR2 and TLR4 was reduced after treatment with L. acidophilus in HT29 cells challenged with SesE, but the downregulation of TLR4 was higher.
- L. acidophilus plays critical roles in regulating TLR4 expression and inducing an anti-inflammatory response via decreasing inflammatory cytokine generation. Similar to our results, decreases in the expression of TLR4 in response to probiotics and Salmonella have been reported by Bermudez-Brito et al [26] in Spain. Villena et al [33] reported that Lactobacillus jensenii TL2937 attenuates the inflammatory response triggered by the activation of TLR4 in intestinal epithelial cells. Our results demonstrated that the use of L. acidophilus for the treatment of cells infected with SesE significantly boosted innate immune responses. Overall, the obtained results suggested that L. acidophilus has strong immunomodulatory properties and can modulate the expression of TLR2 and TLR4 in HT29 intestinal epithelial cells challenged with SesE. These observations support that L. acidophilus may have applications in the prevention and/or treatment of inflammatory diseases. Further in vivo and in vitro investigations are required to elucidate the mechanisms of these anti-inflammatory effects in future studies.
DISCUSSION
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
-
This paper reports part of a research project approved by the Food Microbiology Research Center, Tehran University of Medical Sciences and Health Services (Contract No. 26627).
Article information
- 1. Vandenplas Y, Huys G, Daube G. Probiotics: an update. J Pediatr (Rio J) 2015;91:6−21. https://doi.org/10.1016/j.jped.2014.08.005. PMID: 10.1016/j.jped.2014.08.005. PMID: 25458874.ArticlePubMed
- 2. Thomas CM, Versalovic J. Probiotics-host communication: modulation of signaling pathways in the intestine. Gut Microbes 2010;1:148−63. https://doi.org/10.4161/gmic.1.3.11712. PMID: 10.4161/gmic.1.3.11712. PMID: 20672012.ArticlePubMedPMC
- 3. Hemaiswarya S, Raja R, Ravikumar R, et al. Mechanism of action of probiotics. Braz Arch Biol Technol 2013;56:113−9. https://doi.org/10.1590/S1516-89132013000100015. PMID: 10.1590/S1516-89132013000100015.Article
- 4. Ambalam P, Raman M, Purama RK, et al. Probiotics, prebiotics and colorectal cancer prevention. Best Pract Res Clin Gastroenterol 2016;30:119−31. https://doi.org/10.1016/j.bpg.2016.02.009. PMID: 10.1016/j.bpg.2016.02.009. PMID: 27048903.ArticlePubMed
- 5. Taverniti V, Guglielmetti S. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: proposal of paraprobiotic concept). Genes Nutr 2011;6:261−74. https://doi.org/10.1007/s12263-011-0218-x. PMID: 10.1007/s12263-011-0218-x. PMID: 21499799.ArticlePubMedPMC
- 6. Vidal K, Donnet-Hughes A, Granato D. Lipoteichoic acids from Lactobacillus johnsonii strain La1 and Lactobacillus acidophilus strain La10 antagonize the responsiveness of human intestinal epithelial HT29 cells to lipopolysaccharide and gram-negative bacteria. Infect Immun 2002;70:2057−64. https://doi.org/10.1128/IAI.70.4.2057-2064.2002. PMID: 10.1128/IAI.70.4.2057-2064.2002. PMID: 11895971.ArticlePubMedPMC
- 7. Zou W, Frye JG, Chang CW, et al. Microarray analysis of antimicrobial resistance genes in Salmonella enterica from preharvest poultry environment. J Appl Microbiol 2009;107:906−14. https://doi.org/10.1111/j.1365-2672.2009.04270.x. PMID: 10.1111/j.1365-2672.2009.04270.x. PMID: 19320942.ArticlePubMed
- 8. Eshraghi S, Dalall MMS, Fardsanei F, et al. Salmonella enteritidis and antibiotic resistance patterns: a study on 1950 children with diarrhea. Tehran Univ Med J 2010;67:876−82.
- 9. Rocha TS, Baptista AA, Donato TC, et al. Evaluation of in vitro and in vivo adhesion and immunomodulatory effect of Lactobacillus species strains isolated from chickens. Poult Sci 2012;91:362−9. https://doi.org/10.3382/ps.2011-01803. PMID: 10.3382/ps.2011-01803. PMID: 22252349.ArticlePubMed
- 10. Servin AL. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol Rev 2004;28:405−40. https://doi.org/10.1016/j.femsre.2004.01.003. PMID: 10.1016/j.femsre.2004.01.003. PMID: 15374659.ArticlePubMed
- 11. Feng J, Wang L, Zhou L, et al. Using in vitro immunomodulatory properties of Lactic acid bacteria for selection of probiotics against Salmonella infection in broiler chicks. PLoS One 2016;11:e0147630https://doi.org/10.1371/journal.pone.0147630. PMID: 10.1371/journal.pone.0147630. PMID: 26799658.ArticlePubMedPMC
- 12. Plaza-Diaz J, Gomez-Llorente C, Fontana L, et al. Modulation of immunity and inflammatory gene expression in the gut, in inflammatory diseases of the gut and in the liver by probiotics. World J Gastroenterol 2014;20:15632−49. https://doi.org/10.3748/wjg.v20.i42.15632. PMID: 10.3748/wjg.v20.i42.15632. PMID: 25400447.ArticlePubMedPMC
- 13. Majewska M, Szczepanik M. The role of Toll-like receptors (TLR) in innate and adaptive immune responses and their function in immune response regulation. Postepy Hig Med Dosw (Online) 2006;60:52−63. In Polish. PMID: 16474276.PubMed
- 14. Villena J, Kitazawa H. Modulation of intestinal TLR4-inflammatory signaling pathways by probiotic microorganisms: lessons learned from Lactobacillus jensenii TL2937. Front Immunol 2014;4:512https://doi.org/10.3389/fimmu.2013.00512. PMID: 10.3389/fimmu.2013.00512. PMID: 24459463.ArticlePubMedPMC
- 15. Soltan Dallal MM, Mojarrad M, Baghbani F, et al. Effects of probiotic Lactobacillus acidophilus and Lactobacillus casei on colorectal tumor cells activity (CaCo-2). Arch Iran Med 2015;18:167−72. https://doi.org/0151803/AIM.006. PMID: 25773690.ArticlePubMed
- 16. El-Nezami H, Kankaanpaa P, Salminen S, et al. Ability of dairy strains of lactic acid bacteria to bind a common food carcinogen, aflatoxin B1. Food Chem Toxicol 1998;36:321−6. https://doi.org/10.1016/S0278-6915(97)00160-9. PMID: 10.1016/S0278-6915(97)00160-9. PMID: 9651049.ArticlePubMed
- 17. Pagliaro G, Battino M. The use of probiotics in gastrointestinal diseases. Med J Nutrition Metab 2010;3:105−13. https://doi.org/10.1007/s12349-010-0008-9. PMID: 10.1007/s12349-010-0008-9.Article
- 18. Poppi LB, Rivaldi JD, Coutinho TS, et al. Effect of Lactobacillus sp. isolates supernatant on Escherichia coli O157: H7 enhances the role of organic acids production as a factor for pathogen control. Pesq Vet Bras 2015;35:353−9. https://doi.org/10.1590/S0100-736X2015000400007. PMID: 10.1590/S0100-736X2015000400007.Article
- 19. Khanian SI, Mojgani N, Ahmedi MK. Characterization of partially purified bacteriocin like substance (BLIS) produced by probiotic Lactobacillus strains. Int J Enteric Pathog 2014;2:e17426https://doi.org/10.17795/ijep17426.Article
- 20. Toumi R, Soufli I, Rafa H, et al. Probiotic bacteria lactobacillus and bifidobacterium attenuate inflammation in dextran sulfate sodium-induced experimental colitis in mice. Int J Immunopathol Pharmacol 2014;27:615−27. PMID: 10.1177/039463201402700418. PMID: 25572742.ArticlePubMed
- 21. Wu Z, Pan DD, Guo Y, et al. Structure and anti-inflammatory capacity of peptidoglycan from Lactobacillus acidophilus in RAW-264.7 cells. Carbohydr Polym 2013;96:466−73. https://doi.org/10.1016/j.carbpol.2013.04.028. PMID: 10.1016/j.carbpol.2013.04.028. PMID: 23768588.ArticlePubMed
- 22. Melmed G, Thomas LS, Lee N, et al. Human intestinal epithelial cells are broadly unresponsive to Toll-like receptor 2-dependent bacterial ligands: implications for host-microbial interactions in the gut. J Immunol 2003;170:1406−15. https://doi.org/10.4049/jimmu-nol.170.3.1406. PMID: 10.4049/jimmunol.170.3.1406. PMID: 12538701.ArticlePubMed
- 23. Otte JM, Cario E, Podolsky DK. Mechanisms of cross hyporesponsiveness to Toll-like receptor bacterial ligands in intestinal epithelial cells. Gastroenterology 2004;126:1054−70. https://doi.org/10.1053/j.gastro.2004.01.007. PMID: 10.1053/j.gastro.2004.01.007. PMID: 15057745.ArticlePubMed
- 24. Furrie E, Macfarlane S, Thomson G, et al. Toll-like receptors-2, -3 and -4 expression patterns on human colon and their regulation by mucosal-associated bacteria. Immunology 2005;115:565−74. https://doi.org/10.1111/j.1365-2567.2005.02200.x. PMID: 10.1111/j.1365-2567.2005.02200.x. PMID: 16011525.ArticlePubMedPMC
- 25. Vizoso Pinto MG, Rodriguez Gómez M, Seifert S, et al. Lactobacilli stimulate the innate immune response and modulate the TLR expression of HT29 intestinal epithelial cells in vitro. Int J Food Microbiol 2009;133:86−93. https://doi.org/10.1016/j.ijfoodmicro.2009.05.013. PMID: 10.1016/j.ijfoodmicro.2009.05.013. PMID: 19523707.ArticlePubMed
- 26. Bermudez-Brito M, Muñoz-Quezada S, Gomez-Llorente C, et al. Human intestinal dendritic cells decrease cytokine release against Salmonella infection in the presence of Lactobacillus paracasei upon TLR activation. PLoS One 2012;7:e43197https://doi.org/10.1371/journal.pone.0043197. PMID: 10.1371/journal.pone.0043197. PMID: 22905233.ArticlePubMedPMC
- 27. Vinderola G, Matar C, Perdigon G. Role of intestinal epithelial cells in immune effects mediated by gram-positive probiotic bacteria: involvement of toll-like receptors. Clin Diagn Lab Immunol 2005;12:1075−84. https://doi.org/10.1128/CDLI.12.9.1075-1084.2005. PMID: 16148174.ArticlePubMedPMC
- 28. Castillo NA, Perdigón G, de Moreno de Leblanc A. Oral administration of a probiotic Lactobacillus modulates cytokine production and TLR expression improving the immune response against Salmonella enterica serovar Typhimurium infection in mice. BMC Microbiol 2011;11:177https://doi.org/10.1186/1471-2180-11-177. PMID: 10.1186/1471-2180-11-177. PMID: 21813005.ArticlePubMedPMC
- 29. Finamore A, Roselli M, Imbinto A, et al. Lactobacillus amylovorus inhibits the TLR4 inflammatory signaling triggered by enterotoxigenic Escherichia coli via modulation of the negative regulators and involvement of TLR2 in intestinal Caco-2 cells and pig explants. PLoS One 2014;9:e94891https://doi.org/10.1371/journal.pone.0094891. PMID: 10.1371/journal.pone.0094891. PMID: 24733511.ArticlePubMedPMC
- 30. Cammarota M, De Rosa M, Stellavato A, et al. In vitro evaluation of Lactobacillus plantarum DSMZ 12028 as a probiotic: emphasis on innate immunity. Int J Food Microbiol 2009;135:90−8. https://doi.org/10.1016/j.ijfoodmicro.2009.08.022. PMID: 10.1016/j.ijfoodmicro.2009.08.022. PMID: 19748696.ArticlePubMed
- 31. Wang S, Ng LH, Chow WL, et al. Infant intestinal Enterococcus faecalis down-regulates inflammatory responses in human intestinal cell lines. World J Gastroenterol 2008;14:1067−76. https://doi.org/10.3748/wjg.14.1067. PMID: 10.3748/wjg.14.1067. PMID: 18286689.ArticlePubMedPMC
- 32. Miettinen M, Veckman V, Latvala S, et al. Live Lactobacillus rhamnosus and Streptococcus pyogenes differentially regulate Toll-like receptor (TLR) gene expression in human primary macrophages. J Leukoc Biol 2008;84:1092−100. https://doi.org/10.1189/jlb.1206737. PMID: 10.1189/jlb.1206737. PMID: 18625909.ArticlePubMed
- 33. Villena J, Suzuki R, Fujie H, et al. Immunobiotic Lactobacillus jensenii modulates the Toll-like receptor 4-induced inflammatory response via negative regulation in porcine antigen-presenting cells. Clin Vaccine Immunol 2012;19:1038−53. https://doi.org/10.1128/CVI.00199-12. PMID: 10.1128/CVI.00199-12. PMID: 22573738.ArticlePubMedPMC
REFERENCES


| Primer | Sequence (5′ to 3′) | Reference |
|---|---|---|
| TLR2 F | GCA GAA GCG CTG GGG AAT GG | [17] |
| TLR2 R | GGA TGC CTA CTG GGT GGA GAA | [17] |
| TLR4 F | GGT GGA AGT TGA ACG AAT GG | [17] |
| TLR4 R | CCA GCA AGA AGC ATCAGG TG′ | [17] |
| ACTB F | CTG GAA GAT GGT GAT GGG AT | [9] |
| ACTB R | GGA TTT GGT CGT ATT GGG CG | [9] |
Figure & Data
References
Citations
- Ligilactobacillus salivarius 7247 Strain: Probiotic Properties and Anti-Salmonella Effect with Prebiotics
Vyacheslav M. Abramov, Igor V. Kosarev, Andrey V. Machulin, Evgenia I. Deryusheva, Tatiana V. Priputnevich, Alexander N. Panin, Irina O. Chikileva, Tatiana N. Abashina, Ashot M. Manoyan, Anna A. Ahmetzyanova, Olga E. Ivanova, Tigran T. Papazyan, Ilia N. N
Antibiotics.2023; 12(10): 1535. CrossRef - Anti-Salmonella Defence and Intestinal Homeostatic Maintenance In Vitro of a Consortium Containing Limosilactobacillus fermentum 3872 and Ligilactobacillus salivarius 7247 Strains in Human, Porcine, and Chicken Enterocytes
Vyacheslav M. Abramov, Igor V. Kosarev, Andrey V. Machulin, Evgenia I. Deryusheva, Tatiana V. Priputnevich, Alexander N. Panin, Irina O. Chikileva, Tatiana N. Abashina, Ashot M. Manoyan, Anna A. Akhmetzyanova, Dmitriy A. Blumenkrants, Olga E. Ivanova, Tig
Antibiotics.2023; 13(1): 30. CrossRef - Lactobacillus acidophilus ATCC 4356 Exopolysaccharides Suppresses Mediators of Inflammation through the Inhibition of TLR2/STAT-3/P38-MAPK Pathway in DEN-Induced Hepatocarcinogenesis in Rats
Ola M. S. Khedr, Sawsan M. El-Sonbaty, Fatma S. M. Moawed, Eman I. Kandil, Basma E. Abdel-Maksoud
Nutrition and Cancer.2022; 74(3): 1037. CrossRef - Osmoporation is a versatile technique to encapsulate fisetin using the probiotic bacteria Lactobacillus acidophilus
Eduardo Wagner Vasconcelos de Andrade, Sebastien Dupont, Laurent Beney, Roberta Targino Hoskin, Márcia Regina da Silva Pedrini
Applied Microbiology and Biotechnology.2022; 106(3): 1031. CrossRef - The Game for Three: Salmonella–Host–Microbiota Interaction Models
Krzysztof Grzymajlo
Frontiers in Microbiology.2022;[Epub] CrossRef - The Functional Roles of Lactobacillus acidophilus in Different Physiological and Pathological Processes
Huijuan Gao, Xin Li, Xiatian Chen, Deng Hai, Chuang Wei, Lei Zhang, Peifeng Li
Journal of Microbiology and Biotechnology.2022; 32(10): 1226. CrossRef - Improving bioactive properties of peach juice using Lactobacillus strains fermentation: Antagonistic and anti-adhesion effects, anti-inflammatory and antioxidant properties, and Maillard reaction inhibition
Seyed Mohammad Bagher Hashemi, Dornoush Jafarpour, Mohammad Jouki
Food Chemistry.2021; 365: 130501. CrossRef - The immune regulatory role of Lactobacillus acidophilus: An updated meta-analysis of randomized controlled trials
Wei Zhao, Yangshuo Liu, Lai-Yu Kwok, Tiequan Cai, Wenyi Zhang
Food Bioscience.2020; 36: 100656. CrossRef - Gene expression changes as predictors of the immune-modulatory effects of probiotics: Towards a better understanding of strain-disease specific interactions
Frida Gorreja
NFS Journal.2019; 14-15: 1. CrossRef - A review on anti-adhesion therapies of bacterial diseases
Arezoo Asadi, Shabnam Razavi, Malihe Talebi, Mehrdad Gholami
Infection.2019; 47(1): 13. CrossRef
 PubReader
PubReader Cite
Cite



